0090-9556/05/3306-749–753$20.00DRUG METABOLISM AND DISPOSITION
Copyright 2005 by The American Society for Pharmacology and Experimental Therapeutics
ROLE OF CYP2C9 AND ITS VARIANTS (CYP2C9*3 AND CYP2C9*13) IN THE METABOLISM OF LORNOXICAM IN HUMANS
Yingjie Guo, Yifan Zhang, Ying Wang, Xiaoyan Chen, Dayong Si, Dafang Zhong,
College of Life Science, Jilin University, Changchun, China (Y.G., Y.W., D.S., H.Z.); Laboratory of Drug Metabolism andPharmacokinetics, Shenyang Pharmaceutical University, Shenyang, China (Y.Z., X.C., D.Z.); and School of Pharmacy, Universityof Otago, Dunedin, New Zealand (J.P.F.)
Received January 8, 2005; accepted March 9, 2005
ABSTRACT: CYP2C9 is an important member of the cytochrome P450 enzyme postmitochondrial supernatant (S9) from transfected cells were superfamily with some 12 CYP2C9 alleles (*1-*12) being previously lower than those from wild-type CYP2C9*1. Mean values of K reported. Recently, we identified a new CYP2C9 allele with a V for CYP2C9*1, *3, and *13 were 1.24, 1.61, and 2.79 M and Leu90Pro mutation in a Chinese poor metabolizer of lornoxicam [Si 0.83, 0.28, and 0.22 pmol/min/pmol, respectively. Intrinsic clear- D, Guo Y, Zhang Y, Yang L, Zhou H, and Zhong D (2004) Pharma- ance values (V /K ) for variant CYP2C9*3 and CYP2C9*13 on the cogenetics 14:465–469]. The new allele, designated CYP2C9*13, basis of CYP2C9 protein levels were separately decreased to 28% was found to occur in approximately 2% of the Chinese population. and 12% compared with wild type. In a subsequent clinical study, To examine enzymatic activity of the CYP2C9*13 allele, kinetic the AUC of lornoxicam was increased by 1.9-fold and its oral parameters for lornoxicam 5-hydroxylation were determined in clearance (CL/F) decreased by 44% in three CYP2C9*1/*13 sub- COS-7 cells transiently transfected with pcDNA3.1 plasmids car- jects, compared with CYP2C9*1/*1 individuals. This suggests that rying wild-type CYP2C9*1, variant CYP2C9*3, and CYP2C9*13 the CYP2C9*13 allele is associated with decreased enzymatic ac- cDNA. The protein levels of cDNA-expressed CYP2C9*3 and *13 in tivity both in vitro and in vivo.
CYP2C9 constitutes approximately 20% of the cytochrome P450
show decreased enzymatic activity in vitro and in vivo (Crespi et al.,
protein content of human liver microsomes and is responsible for the
1997; Aithal et al., 1999; Imai et al., 2000; Dickmann et al., 2001;
metabolism of many clinically important drugs. These include drugs
Kidd et al., 2001; Kirchheiner et al., 2002; Allabi et al., 2004;
with a narrow therapeutic index such as warfarin and phenytoin and
other routinely prescribed drugs such as acenocoumarol, tolbutamide,
Recently, a new CYP2C9 allele designated CYP2C9*13 has been
losartan, glipizide, and some nonsteroidal anti-inflammatory drugs
identified in a Chinese poor metabolizer of lornoxicam. The allele
possesses a T269C transversion in exon 2 of CYP2C9 that leads to a
The CYP2C9 gene is highly polymorphic. At least 13 CYP2C9
Leu90Pro substitution. Frequency analysis shows that approximately
alleles have been identified to date and most of them are associated
2% of the Chinese populations carry the allele (Si et al., 2004).
with reduced CYP2C9 activity. Among them, CYP2C9*3, with an
Genotyping of this poor lornoxicam metabolizer revealed a
Ile359Leu mutation, has been most widely studied. In vitro studies
CYP2C9*3/*13 genotype with the two mutations located on separate
show it has significantly impaired catalytic activity to various
alleles. His lornoxicam half-life of about 105 h was markedly longer
CYP2C9 substrates relative to the wild type (Takanashi et al., 2000).
than that of other CYP2C9*1/*3 and CYP2C9*1/*1 carriers (half-lives
In vivo investigations show that individuals heterozygous and ho-
of 5.8 – 8.1 and 3.2– 6.3 h, respectively; Zhang et al., 2005), suggest-
mozygous for CYP2C9*3 have reduced intrinsic clearance of warfa-
ing that the CYP2C9*13 allele has a larger effect on CYP2C9 meta-
rin, phenytoin, and glipizide and are more at risk of clinical toxicity
bolic capability than other alleles.
from these drugs (Aithal et al., 1999; Kidd et al., 1999; Ninomiya et
CYP2C9 has been shown to be the primary enzyme responsible for
al., 2000). Other CYP2C9 alleles such as CYP2C9*2 (Arg144Cys),
the biotransformation of the nonsteroidal anti-inflammatory drug lor-
CYP2C9*4 (Ile359Thr), CYP2C9*5 (Asp360Glu), CYP2C9*6 (null
noxicam to its major metabolite, 5Ј-hydroxylornoxicam, in human
allele), CYP2C9*11 (Arg335Trp), and CYP2C9*12 (Pro489Ser) also
liver microsomes (Bonnabry et al., 1996; Kohl et al., 2000). Recently,it was reported that lornoxicam 5Ј-hydroxylation by the CYP2C9*3allele was markedly reduced compared with wild type, both in vitro
Project supported by the National Natural Science Foundation of China, No.
and in vivo (Iida et al., 2004; Zhang et al., 2005). Thus, lornoxicam is
an ideal substrate for the study of CYP2C9 enzyme activity. The
purpose of this study was to compare the enzymatic activity of
CYP2C9*1, CYP2C9*3, and CYP2C9*13 toward lornoxicam both in
ABBREVIATIONS: DMEM, Dulbecco’s modified Eagle’s medium; P450, cytochrome P450; BCIP/NBT, 5-bromo-4-chloro-3-indolyl phosphate/ nitro blue tetrazolium; CPR, NADPH-cytochrome P450 reductase; AUC, area under the plasma concentration-time curve; CL/F, oral clearance.
vitro in appropriately transfected COS-7 cells and in vivo in subjects
(Invitrogen) as a standard with ImageJ software (National Institutes of Health,
with CYP2C9*1/*3, CYP2C9*1/*13, and CYP2C9*1/*1 genotypes. In Vitro Lornoxicam 5-Hydroxylation of Recombinant CYP2C9 Pro- tein. Lornoxicam 5Ј-hydroxylation activity of the recombinant CYP2C9 pro- Materials and Methods
tein was determined as described previously (Kohl et al., 2000) with minor
Materials. Lornoxicam was purchased from Shangdi Xinshiji Medical
modifications. S9 fraction containing CYP2C9s was incubated with 100 mM
Academy (Beijing, China.). 5Ј-Hydroxylornoxicam was provided by the Lab-
Tris buffer (pH 7.5), 200 M NADPH, and lornoxicam at 37°C for 1 h in the
oratory of Microorganisms, Shenyang Pharmaceutical University (Shenyang,
presence or absence of CPR. The reaction was stopped by addition of 500 l
China). Dulbecco’s modified Eagle’s medium (DMEM), pcDNA3.1(ϩ), and
of methanol and stored overnight at Ϫ20°C to allow complete protein precip-
LipofectAMINE 2000 were purchased from Invitrogen (Carlsbad, CA). Fetal
itation. After centrifugation for 30 min at 12,000 rpm, the supernatants were
bovine serum was purchased from Tianjin, H&Y Bio Co. Ltd. (Tianjing,
reduced to 100 l by warming at 65°C and subjected to high performance
China). KpnI, XhoI, and DpnI enzymes were purchased from New England
liquid chromatography assay. High performance liquid chromatography was
Biolabs (Beverly, MA). Pfu DNA polymerase was purchased from Bio Basic
Inc. (Toronto, ON, Canada). A pREP9 plasmid containing human CYP2C9*1
Technologies, Palo Alto, CA) using a mobile phase of 0.1 M NaH PO , pH
cDNA and E. coli Top 10 were provided by the Department of Pathophysiol-
6.0/acetonitrile (7:3), at a flow rate of 1 ml/min. Detection was by UV
ogy and the Laboratory of Medical Molecular Biology, School of Medicine,
absorption at 371 nm. Under these conditions, retention times of 5Ј-hydroxy-
Zhejiang University (Zhejiang, China). Rabbit anti-human cytochrome P450
lornoxicam and lornoxicam were 6.9 and 11.8 min, respectively. A six-point
2C9 antibody was purchased from Serotec (Oxford, UK). Alkaline phos-
standard curve was used to quantify 5Ј-hydroxylornoxicam.
phatase-labeled anti-rabbit IgG, 5-bromo-4-chloro-3-indolyl phosphate/nitro
In Vivo Lornoxicam Metabolism. The study was approved by the Inde-
blue tetrazolium (BCIP/NBT) and bovine serum albumin were purchased from
pendent Ethics Committee of the People’s Hospital of Liaoning Province
Beijing Dingguo Biotechnology Development Center (Beijing, China).
(Shenyang, China). Genotyping of CYP2C9*3 and CYP2C9*13 was carried
NADPH was purchased from Roche Molecular Biochemicals (Basel, Switzer-
out as described previously (Si et al., 2004; Zhang et al., 2005). Thirteen
land). COS-7 cells were kindly donated by the Vaccination Center, Jilin
CYP2C9*1 homozygotes, 7 CYP2C9*3 heterozygotes, and 3 CYP2C9*13
University (Changchun, China). Human recombinant NADPH-P450 reductase
heterozygotes participated in the phenotyping study. All subjects were in good
(CPR) was purchased from MBL International Corporation (Woburn, MA).
health and were required to refrain from all medication and alcohol prior to the
All other reagents were of analytical grade.
pharmacokinetic study. In vivo lornoxicam metabolism was performed accord-
Construction of Expression Plasmids. CYP2C9*1 cDNA in pREP9 plas-
ing to the method described previously (Zhang et al., 2005). In brief, after a
mid was subcloned into pcDNA3.1(ϩ) by digestion with KpnI and XhoI
single oral dose of 8 mg of lornoxicam, blood samples were collected before
enzymes. Site-directed mutagenesis to introduce the A3 C transition at posi-
dosing, and at 0.5, 1, 1.5, 2, 2.5, 3, 4, 6, 8, 10, 13, and 24 h postdose. Plasma
tion 1075 (CYP2C9*3) and the T3 C transition at position 269 (CYP2C9*13)
concentrations of lornoxicam were determined using a validated liquid chro-
was performed using pcDNA3.1(ϩ) plasmids carrying CYP2C9*1 cDNA as
matography-tandem mass spectrometry method reported elsewhere (Zeng et
the template for polymerase chain reaction amplification by Pfu DNA poly-
merase. The specific base transition was introduced into the amplificationproducts by a pair of completely complementary primers containing substi-tuted base. The mutagenic primers for CYP2C9*3 and *13 were 5Ј-CGAGG-TCCAGAGATACCTTGACCTTCTCCCCAC-3Ј and 5Ј-GGAAGCCCTGA-TTGATCCTGGAGAGGAGTTTTCTG-3Ј, respectively (mutations und-erlined). After incubation with DpnI enzyme, the origin templates were dig-ested, but the new amplified polymerase chain reaction products containingsubstituted base remained and were transformed to E. coli Top 10. Clonescontaining the desired nucleotide change were identified by sequencing carriedout by Shanghai Sangon Biological Engineering Technology and Service Co. Ltd. (Shanghai, China). Transfection of COS-7 Cells and Preparation of Postmitochondrial Supernatant (S9). COS-7 cells were seeded into 10-cm culture flasks in DMEM containing 10% fetal bovine serum, 100 U/ml penicillin, and 100 g/ml streptomycin. When cells were 90 to 95% confluent, the culture medium was replaced with DMEM without penicillin and streptomycin, and the CYP2C9 expression plasmids (24 g/flask), purified with a QIAGEN plasmid mini kit (QIAGEN, Valencia, CA), were transfected into COS-7 cells using LipofectAMINE 2000 at 60 l/flask, as per the manufacturer’s instructions. Forty-eight hours after transfection, cells were scraped from the culture flask and washed twice with Ca2ϩ- and Mg2ϩ-free Hanks’ solution. The pellets were resuspended in 20 mM potassium phosphate buffer, pH 7.4, containing 0.2 mM EDTA, 1 mM dithiothreitol, and 20% glycerol, and sonicated with twelve 5-s pulses at 23% power of a Sonics Vibra-Cell sonicator (Sonics & Materials, Inc., Newtown, CT). The homogenate was centrifuged at 9000g, 4°C for 20 min and the postmitochondrial supernatant (S9 fraction) collected for assay or storage at Ϫ70°C. Protein concentrations in S9 were determined by the Bradford method (Bradford, 1976) using bovine serum albumin as standard. Quantification of CYP2C9 Protein by Western Blotting. S9 fraction (50
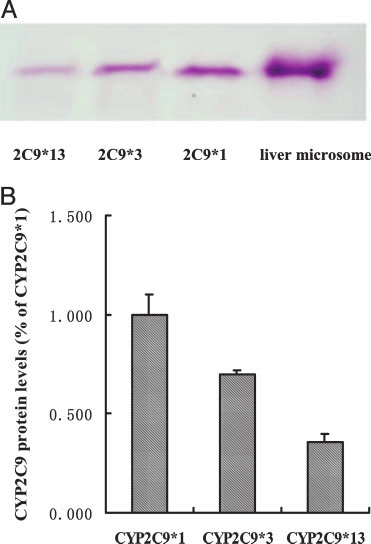
FIG. 1. CYP2C9 protein levels in postmitochondrial supernatant (S9) from COS-7
g) and human liver microsomes (10 g) were separated on 10% sodium
cells expressing wild-type and variant CYP2C9s. A, immunoblot analysis of re-
dodecyl sulfate-polyacrylamide gels and transferred to a polyvinylidene diflu-
combinant human CYP2C9 protein. S9 fraction (50 g) isolated from COS-7 cells
oride membrane (Millipore Corporation, Billerica, MA). The membrane was
transfected with the wild-type and variant CYP2C9 cDNA was utilized for immu-noblotting using anti-human CYP2C9 antibody. Human liver microsome (10 g)
incubated with rabbit anti-human cytochrome P450 2C9 antibody as the
was used as positive control. B, protein levels of CYP2C9 were quantified by
primary antibody and then with alkaline phosphatase-labeled anti-rabbit IgG as
densitometric analysis. The results are expressed as a percentage of the level of
the secondary antibody. Bands were visualized by incubation with BCIP/NBT
CYP2C9*1. Each bar represents the mean Ϯ S.E.M. of three independent experi-
and quantified by microsomes from insect cells expressing human CYP2C9
CYP2C9 VARIANTS AND LORNOXICAM METABOLISM
FIG. 2. Michaelis-Menten kinetics of lornoxicam by postmitochondrial supernatant(S9) from COS-7 cells expressing wild-type and variant CYP2C9s. S9 fractioncorresponding to 500 g of protein was incubated with different concentrations oflornoxicam in the absence of CPR. Experimental conditions are described underMaterials and Methods. Each point represents the mean of three independentexperiments. f, CYP2C9*1; Œ, CYP2C9*3; , CYP2C9*13.
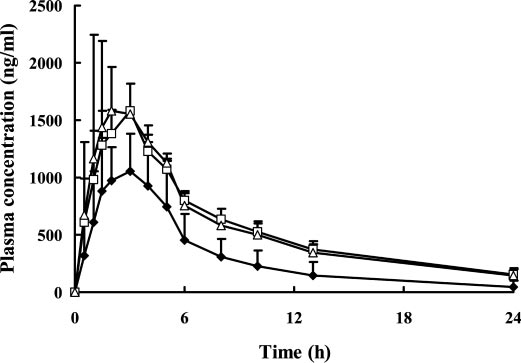
FIG. 3. Plasma concentration-time curves of lornoxicam in healthy subjects withgenotype CYP2C9*1/*1 (ࡗ) (n ϭ 13), CYP2C9*1/*3 (Ⅺ) (n ϭ 7), and CYP2C9*1/Data Analysis. Michaelis-Menten analysis was performed by nonlinear *13 (‚) (n ϭ 3) after a single oral dose of 8 mg of lornoxicam.
regression curve fitting using the computer program Prism v4.0 (GraphPadSoftware Inc., San Diego, CA). Pharmacokinetic parameters were calculated
matic activities were normalized to CYP2C9 protein levels, intrinsic
using standard noncompartmental methods. Student’s t test was used for
/K ) for variant CYP2C9*3 and CYP2C9*13
intergroup comparison. A value of P Ͻ 0.05 was considered to be statistically
were also separately decreased to 28% and 12% compared with wild
type. Clearly, the activity of CYP2C9*13 is lower than that of
In the in vivo study, the CYP2C9 genotype significantly affected
A representative immunoblot of S9 fraction prepared from COS-7
the pharmacokinetics of lornoxicam (Fig. 3; Table 2). The AUC of
cells expressing CYP2C9*1, CYP2C9*3, and CYP2C9*13 proteins is
lornoxicam increased 1.9-fold and CL/F was decreased by 44% in
presented in Fig. 1. All constructs yielded immunodetectable CYP2C9
CYP2C9*1/*13 individuals compared with CYP2C9*1/*1 individu-
protein, as did human liver microsomes. The expressed protein level
als. Similar changes were found in CYP2C9*1/*3 individuals.
of CYP2C9*1 was 7.51 pmol/mg S9 protein, and the expression levelsof variant CYP2C9*3 and *13 were 69.9% and 35.5%, respectively,of that of CYP2C9*1. Discussion
The effect of exogenous CPR on lornoxicam 5Ј-hydroxylation was
To investigate the catalytic activity of the CYP2C9*13 allele in
studied by incubation with recombinant CYP2C9*1. S9 fraction con-
vitro, we established a COS cell expression system. This has been
taining 3.75 pmol of CYP2C9*1 was mixed with varying amounts of
widely applied for functional characterization of P450 alleles contain-
CPR and 10 M lornoxicam. When CPR was 0, 5, 12.5, or 25 M, the
ing CYP2C9, CYP2D6, CYP2B6, and CYP2E1 (Veronese et al.,
for CYP2C9*1 was 0.66, 0.74, 0.68, or 0.68
1993; Hu et al., 1997; Marcucci et al., 2002; Jinno et al., 2003).
pmol/min/pmol, respectively. There was no significant alteration in
However, due to the low levels of expression in COS-7 cells, we
catalytic efficiency of CYP2C9*1 with the increasing CPR concen-
failed to quantify CYP2C9 holoenzyme contents by CO-difference
tration, indicating that endogenous reductase is enough for lornoxicam
spectroscopy. Thus, CYP2C9 proteins were quantified by immuno-
metabolism in the COS-7 expression system. Thus, exogenous CPR
blotting with microsomes from insect cells expressing human
was not utilized in the following kinetic study.
CYP2C9 as standard. This kind of method quantifying P450 was also
Michaelis-Menten kinetics of lornoxicam for wild-type and mutant
used for the functional characterization of CYP2D6, CYP2B6, and
CYP2C9 is shown in Fig. 2. Corresponding kinetic parameters are
CYP2E1 allelic variants in the COS expression system (Marcucci et
summarized in Table 1. On the basis of protein levels of S9 fraction,
al., 2002; Hanioka et al., 2003; Jinno et al., 2003). Our results show
both CYP2C9*3 and CYP2C9*13 exhibited lower intrinsic clearance
that COS-7 cells can efficiently express active CYP2C9 protein. The
of lornoxicam (P Ͻ 0.01) than did wild-type CYP2C9*1 resulting
protein levels of cDNA-expressed CYP2C9*3 and *13 in S9 fraction
(1.3-fold and 2.3-fold, respectively) and
from COS-7 cells were lower than those in wild-type CYP2C9*1. The
(76% and 90%, respectively). When the enzy-
reduced protein levels in the CYP2C9 variants may contribute to
Kinetic parameters for lornoxicam 5Ј-hydroxylation from COS-7 cells expressing wild-type and variant CYP2C9s
Each value represents the mean Ϯ S.E.M. of three independent experiments.
** P Ͻ 0.01 versus CYP2C9*1. Pharmacokinetic parameters of lornoxicam in healthy subjects with CYP2C9*1/*1, CYP2C9*1/*3, and CYP2C9*1/*13 genotype
Data are given as mean and 95% confidence interval (in parentheses).
* P Ͻ 0.05, **P Ͻ 0.01 versus CYP2C9*1/*1.
lower transcription, translation efficiency, and protein stability. Oth-
Our results show that the activities of both CYP2C9*3 and
erwise, although polyclonal antibody was used, the immunoreactivity
CYP2C9*13 toward lornoxicam in vitro are compatible with their
of mutant CYP2C9s may be altered by mutagenization. Therefore, the
activities in vivo, and there is a reasonable correlation between in vitro
enzymatic activities were assayed in two ways, on the basis of S9
activity and in vivo metabolic clearance of lornoxicam. Recently, it
protein level and CYP2C9 protein level. These data obtained using the
was reported that individuals carrying the CYP2C9*3 allele are at risk
COS expression system need to be confirmed by a baculovirus sys-
of experiencing drug toxicity, especially of drugs with a narrow
tem, which is better suited to obtain the quantity of P450 by spectral
therapeutic index such as warfarin and phenytoin (Aithal et al., 1999;
analysis. These studies are currently under investigation in our labo-
Kidd et al., 1999; Ninomiya et al., 2000). By extrapolation, one may
speculate that carriers of the CYP2C9*13 allele would experience
In this study, the presence of the CYP2C9*3 allele impairs both
greater risk from these drugs. Therefore, genotyping for CYP2C9*13
intrinsic clearance and systemic clearance of lornoxicam. A recent
may be important to allow individualization of dosing for CYP2C9
report shows that CYP2C9*1/*3 individuals have a 55% decrease in
CL/F and a 1.9-fold increase in AUC of lornoxicam compared with
In conclusion, the Leu90Pro substitution of CYP2C9*13 markedly
CYP2C9*1/*1 individuals (Zhang et al., 2005). The magnitude of
decreases the intrinsic clearance of lornoxicam in vitro and in vivo.
these changes in pharmacokinetic parameters is consistent with our in
The reduction in activity due to CYP2C9*13 is greater than that due
vivo results. Iida et al. (2004) reported that CYP2C9*3 expressed in
to CYP2C9*3 in vitro. Whether carriers of the CYP2C9*13 allele may
baculovirus-infected insect cells significantly decreased lornoxicam
be at greater risk of toxicity from CYP2C9 substrate drugs with a
5Ј-hydroxylation relative to wild type with a 2.3-fold increase in K
narrow therapeutic index remains to be confirmed by further in vivo
(P Ͻ 0.05) and 76% decrease in V
showed a 1.3-fold increase (P Ͼ 0.05) and the V
on the basis of CYP2C9 protein level relative to wild type. The reason
References
for the discrepancy is the difference in the heterologous cell expres-
Aithal GP, Day CP, Kesteven PJL, and Daly AK (1999) Association of polymorphisms in the
sion system. The addition of exogenous reductase did not significantly
cytochrome P450 CYP2C9 with warfarin dose requirement and risk of bleeding complications. Lancet 353:717–719.
affect the lornoxicam 5Ј-hydroxylation by CYP2C9*1, indicating that
Allabi AC, Gala JL, Horsmans Y, Babaoglu MO, Bozkurt A, Heusterspreute M, and Yasar U
the endogenous reductase in COS-7 cells is not limiting for lornoxi-
(2004) Functional impact of CYP2C95, CYP2C96, CYP2C98 and CYP2C911 in vivo among black Africans. Clin Pharmacol Ther 76:113–118.
Blaisdell J, Jorge-Nebert LF, Coulter S, Ferguson SS, Lee SJ, Chanas B, Xi T, Mohrenweiser H,
In addition to the CYP2C9*3 variant, the catalytic activity of a
Ghanayem B, and Goldstein JA (2004) Discovery of new potentially defective alleles of human CYP2C9. Pharmacogenetics 14:527–537.
recently identified CYP2C9*13 variant that contains a Leu90Pro
Bonnabry P, Leemann T, and Dayer P (1996) Role of human liver microsomal CYP2C9 in the
substitution was investigated in this study. Compared with wild-type
biotransformation of lornoxicam. Eur J Clin Pharmacol 49:305–308.
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities
CYP2C9*1, the CYP2C9*13 variant also has lower intrinsic clearance
of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248 –254.
for lornoxicam 5Ј-hydroxylation due to a 2.3-fold increase in K and
Crespi CL and Miller VP (1997) The R144C change in the CYP2C9*2 allele alters interaction
on the basis of CYP2C9 protein level. The
of the cytochrome P450 with NADPH:cytochrome P450 oxidoreductase. Pharmacogenetics7:203–210.
results are consistent with our in vivo observation that individuals
Dickmann LJ, Rettie AE, Kneller MB, Kim RB, Wood AJ, Stein CM, Wilkinson GR, and
with CYP2C9*1/*13 genotype have an impaired clearance of lornoxi-
Schwarz UI (2001) Identification and functional characterization of a new CYP2C9 variant (CYP2C9*5) expressed among African Americans. Mol Pharmacol 60:382–387.
cam compared with individuals with CYP2C9*1/*1 genotype. Inter-
Hanioka N, Tanaka-Kagawa T, Miyata Y, Matsushima E, Makino Y, Ohno A, Yoda R, Jinno H,
estingly, in our study, individuals with CYP2C9*1/*3 and CYP2C9*1/
and Ando M (2003) Functional characterization of three human cytochrome P450 2E1 variants with amino acid substitutions. Xenobiotica 33:575–586.
*13 genotypes reveal the same extent of reduction in oral clearance of
Hu Y, Oscarson M, Johansson I, Yue QY, Dahl ML, Tabone M, Arinco S, Albano E, and
lornoxicam despite the fact that, in vitro, CYP2C9*13 is associated
Ingelman-Sundberg M (1997) Genetic polymorphism of human CYP2E1: characterization of two variant alleles. Mol Pharmacol 51:370 –376.
with a lower intrinsic clearance of lornoxicam than is CYP2C9*3.
Iida I, Miyata A, Arai M, Hirota M, Akimoto M, Higuchi S, Kobayashi K, and Chiba K (2004)
Given the small number of CYP2C9*1/*13 subjects studied (n ϭ
Catalytic roles of CYP2C9 and its variants (CYP2C9*2 and CYP2C9*3) in lornoxicam 5Ј-hydroxylation. Drug Metab Dispos 32:7–9.
3), and in the absence of any individuals homozygous for the
Imai J, Ieiri I, Mamiya K, Miyahara S, Furuumi H, Nanba E, Yamane M, Fukumaki Y, Ninomiya
CYP2C9*13 allele, we recognize that further in vivo studies are
H, Tashiro N, et al. (2000) Polymorphism of the cytochrome P450 (CYP) 2C9 gene in Japanese epileptic patients: genetic analysis of the CYP2C9 locus. Pharmacogenetics 10:85–
required to draw firm conclusions about the role of the CYP2C9*13
allele. According to a crystal structure of CYP2C9 published by
Jinno H, Tanaka-Kagawa T, Ohno A, Makino Y, Matsushima E, Hanioka N, and Ando M (2003)
Functional characterization of cytochrome P450 2B6 allelic variants. Drug Metab Dispos
Williams et al. (2003) and Wester et al., (2004), Leu90 is located in
31:398 – 403.
the B-BЈ loop, which is not the heme-binding region and far from the
Kidd RS, Curry TB, Gallagher S, Edeki T, Blaisdell J, and Goldstein JA (2001) Identification of
binding pocket of substrate. Thus, the reason for the increase in K for
a null allele of CYP2C9 in an African-American exhibiting toxicity to phenytoin. Pharma-cogenetics 11:803– 808.
lornoxicam 5Ј-hydroxylation is not clear. Homology modeling based
Kidd RS, Straughn AB, Meyer MC, Blaisdell J, Goldstein JA, and Dalton JT (1999) Pharma-
on the crystal structure of human CYP2C9 is ongoing in our labora-
cokinetics of chlorpheniramine, phenytoin, glipizide and nifedipine in an individual homozy- gous for the CYP2C9*3 allele. Pharmacogenetics 9:71– 80.
Kirchheiner J, Bauer S, Meineke I, Rohde W, Prang V, Meisel C, Roots I, and Brockmoller J
CYP2C9 VARIANTS AND LORNOXICAM METABOLISM
(2002) Impact of CYP2C9 and CYP2C19 polymorphisms on tolbutamide kinetics and the
Veronese ME, Doecke CJ, Mackenzie PI, McManus ME, Miners JO, Rees DL, Gasser R, Meyer
insulin and glucose response in healthy volunteers. Pharmacogenetics 12:101–109.
UA, and Birkett DJ (1993) Site-directed mutation studies of human liver cytochrome P-450
Kohl C and Steinkellner M (2000) Prediction of pharmacokinetic drug/drug interactions from in
isoenzymes in the CYP2C subfamily. Biochem J 289:533–538.
vitro data: interactions of the nonsteroidal anti-inflammatory drug lornoxicam with oral
Wester MR, Yano JK, Schoch GA, Yang C, Griffin KJ, Stout CD, and Johnson EF (2004) The
anticoagulants. Drug Metab Dispos 28:161–168.
structure of human cytochrome P450 2C9 complexed with flurbiprofen at 2.0-A resolution.
Lee CR, Goldstein JA, and Pieper JA (2002) Cytochrome P450 2C9 polymorphisms: a compre-
J Biol Chem 279:35630 –35637.
hensive review of the in-vitro and human data. Pharmacogenetics 12:251–263.
Williams PA, Cosme J, Ward A, Angove HC, Matak Vinkovic D, and Jhoti H (2003) Crystal
Marcucci KA, Pearce RE, Crespi C, Steimel DT, Leeder JS, and Gaedigk A (2002) Character-
structure of human cytochrome P450 2C9 with bound warfarin. Nature (Lond) 424:464 – 468.
ization of cytochrome P450 2D6.1 (CYP2D6.1), CYP2D6.2, and CYP2D6.17 activities toward
Zhang Y, Zhong D, Si D, Guo Y, Chen X, and Zhou H (2005) Lornoxicam pharmacokinetics in
model CYP2D6 substrates dextromethorphan, bufuralol, and debrisoquine. Drug Metab Dis-
relation to cytochrome P450 2C9 genotype. Br J Clin Pharmacol 59:14 –17. pos 30:595– 601.
Zeng Y, Chen X, Zhang Y, and Zhong D (2004) Determination of lornoxicam in human plasma
Ninomiya H, Mamiya K, Matsuo S, Ieiri I, Higuchi S, and Tashiro N (2000) Genetic polymor-
by LC/MS/MS. Acta Pharmacol Sin 39:132–135.
phism of the CYP2C subfamily and excessive serum phenytoin concentration with central nervous system intoxication. Ther Drug Monit 22:230 –232.
Si D, Guo Y, Zhang Y, Yang L, Zhou H, and Zhong D (2004) Identification of a novel variant
Address correspondence to: Hui Zhou, College of Life Science, Jilin
CYP2C9 allele in Chinese. Pharmacogenetics 14:465– 469.
University, No.115 Jiefang Road, Changchun, 130023, China. E-mail: zhouhui@
Takanashi K, Tainaka H, Kobayashi K, Yasumori T, Hosakawa M, and Chiba K (2000) CYP2C9
Ile359 and Leu359 variants: enzyme kinetic study with seven substrates. Pharmacogenetics10:95–104.
Why do we lose hair? Hair loss affects most people and their emotional response will range from mild concern to traumatic despair. Hair loss also has a time frame whereby it may be a temporary state (eg. from weight loss) or permanent state (eg. male pattern hair loss). Genetically patterned hair loss is also known as male and female pattern baldness . Male and female pattern hair
Palestine's economy is like a coiled spring, says British financier - Business News, Bu. Page 1 of 3 Yasmin Alibhai-Brown : Don't Israel's nuclear weapons count? Palestine's economy is like a coiled spring, says British financier Investment conference backed by Tony Blair told that political progress could spark rapid growth SPONSORED LINKS: Ads by Google QROPS Expert Advice Guide Exp
 vitro in appropriately transfected COS-7 cells and in vivo in subjects
(Invitrogen) as a standard with ImageJ software (National Institutes of Health,
with CYP2C9*1/*3, CYP2C9*1/*13, and CYP2C9*1/*1 genotypes.
vitro in appropriately transfected COS-7 cells and in vivo in subjects
(Invitrogen) as a standard with ImageJ software (National Institutes of Health,
with CYP2C9*1/*3, CYP2C9*1/*13, and CYP2C9*1/*1 genotypes.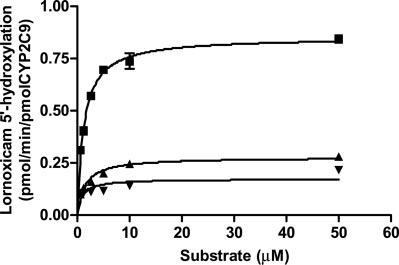
 CYP2C9 VARIANTS AND LORNOXICAM METABOLISM
FIG. 2. Michaelis-Menten kinetics of lornoxicam by postmitochondrial supernatant(S9) from COS-7 cells expressing wild-type and variant CYP2C9s. S9 fractioncorresponding to 500 g of protein was incubated with different concentrations oflornoxicam in the absence of CPR. Experimental conditions are described underMaterials and Methods. Each point represents the mean of three independentexperiments. f, CYP2C9*1; Œ, CYP2C9*3; , CYP2C9*13.
CYP2C9 VARIANTS AND LORNOXICAM METABOLISM
FIG. 2. Michaelis-Menten kinetics of lornoxicam by postmitochondrial supernatant(S9) from COS-7 cells expressing wild-type and variant CYP2C9s. S9 fractioncorresponding to 500 g of protein was incubated with different concentrations oflornoxicam in the absence of CPR. Experimental conditions are described underMaterials and Methods. Each point represents the mean of three independentexperiments. f, CYP2C9*1; Œ, CYP2C9*3; , CYP2C9*13.