International Journal for Parasitology 29 (1999) 619±625
The eect of chloroquine treatment on the infectivity of
Institute of Cell, Animal and Population Biology, Division of Biological Sciences, University of Edinburgh, Edinburgh EH9 3JT, UK
Received 19 October 1998; received in revised form 3 December 1998; accepted 3 December 1998
The antimalarial drug chloroquine has been reported to increase the infectivity of the forms of blood-stage
malaria parasites (gametocytes) that are capable of infecting mosquito vectors. This eect has been demonstrated
convincingly in the short term (12 h post treatment), although several authors have suggested infectivity
enhancement a week or more after treatment. We carried out experiments to investigate the eects of chloroquine
on the longer-term infectivity of gametocytes of the rodent malaria parasite, Plasmodium chabaudi, to Anopheles
stephensi mosquitoes. Gametocytes of chloroquine-treated infections were signi®cantly more infectious than
untreated infections 6 and 7 days post-treatment, although not on days 8 and 9. However, this eect was most likely
the result of a reduction in infectivity in untreated infections, caused by immune activity which was not so
pronounced in chloroquine-treated infections. Gametocytaemia (gametocytes per r.b.c.) showed a strong positive
and linear relationship with infectivity. Infectivity was not in¯uenced by either asexual parasitaemia, asexual density
or anaemia. Parsimonious interpretations of the eect of chloroquine on gametocyte infectivity are discussed.
# 1999 Australian Society for Parasitology Inc. Published by Elsevier Science Ltd. All rights reserved.
Keywords: Anopheles stephensi; Chloroquine; Gametocyte; Infectivity; Malaria; Plasmodium chabaudi; Transmission
the transmission stages of a Plasmodium infec-
failed to signi®cantly reduce malaria prevalence,
tion, gametocytes [2, 3]. However, two other
and resistance to many antimalarial drugs has
spread at an alarming rate [1]. This is partly
enhance transmission have been suggested. First,
because some of the commonly used antimalarial
chemotherapy may induce greater gametocyte
drugs have little parasite killing activity against
production, both through genotypic [4±6] and
short-term phenotypic changes [7]. Given the
generally positive correlation between gametocyte
densities and infectivity to mosquitoes [8, 9], such
* Corresponding author. Present address: Department of
changes are likely to increase transmission.
Plant Sciences, University of Oxford, South Parks Road,
Second, the antimalarial drug chloroquine (CQ)
Oxford OX1 3RB, UK. Tel: 01865-275106; Fax: 01865-
275074; e-mail: angus.buckling@plant-sciences.oxford.ac.uk
has been reported to increase the infectiousness
0020-7519/99/$20.00 # 1999 Australian Society for Parasitology Inc. Published by Elsevier Science Ltd. All rights reserved.
A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
of gametocytes, independent of gametocyte
cyte infectivity with increasing eciency for
21 days following treatment'', using cultured
Data on infectivity enhancement of gameto-
gametocytes of P. falciparum and of the rodent
cytes by CQ are ambiguous. It is clear that CQ
malaria, Plasmodium berghei. They report a sig-
per se does not increase gametocyte infectivity:
ni®cant increase in infectivity through time fol-
the addition of CQ to cultures prior to feeding
mosquitoes did not increase either the pro-
gametocytes, and a non-signi®cant increase with
portions of mosquitoes infected, or the number
P. falciparum gametocytes, although the validity
of these analyses is questionable. Infectivity of
developed [1, 10, 11]. Enhanced gametocyte infec-
gametocytes obtained from naturally infected
tivity 12-h post sub-curative CQ treatment has
CQ-treated volunteers was not signi®cantly
been demonstrated in vivo for the rodent malaria
greater when mixed with the patients' own (CQ-
containing) plasma compared with European
Plasmodium yoelii nigeriensis [14]. However, there
control plasma [17]. A ®eld study investigating
was no reported increase in the infectivity of
factors in¯uencing human infectivity to mosqui-
Plasmodium falciparum gametocytes when mixed
toes also found no P. falciparum infectivity-
with sera collected 24 h after CQ had been taken,
compared with sera before treatment [15].
It is clearly important to determine whether
Similarly, there was no infectivity enhancement
CQ enhances gametocyte infectivity for more
than a matter of hours. We therefore carried out
between 0 and 72 h post-treatment [3].
experiments to investigate gametocyte infectivity
of the rodent malaria, Plasmodium chabaudi, in
enhance per gametocyte infectivity a week or
vivo 6±9 days after CQ treatment, at the time of
more after treatment, but here the data are even
more ambiguous. Curative CQ-treatment of P.
Details of methodology are given in Buckling
falciparum-infected humans resulted in greater
et al. [7]. Male C57/BL/6J mice were infected by
median parasite burdens in Anopheles balabacen-
i.p. inoculation with 106 parasites on day 0.
sis mosquitoes (but not in Anopheles dirus) fed 2
Parasite clones and sample sizes are shown in
and 7 days after treatment, compared with that
Table 1. On day 5 or 6 p.i. (1±2 days before the
before treatment. However, mean oocyst burdens
average peak asexual parasite density), half the
and the proportions of mosquitoes infected did
mice were orally treated with a sub-curative dose
not change [16]. Hogh et al. [17] recently reported
of chloroquine sulphate (12 mg kgÀ1); the others
that ``serum from chloroquine-treated, unin-
received distilled water (controls). In each exper-
fected, non-immune volunteers enhanced gameto-
iment, half the control and half the CQ-treated
a One mouse died from severe anaemia on day 8 post-infection.
A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
mice were picked randomly and fed to H40 4±5-
day-old Anopheles stephensi on day 12 p.i. (except
in the control group in experiment 1, where two
animals out of ®ve were fed to mosquitoes). The
other mice were fed to mosquitoes on day 14 p.i.
Days 12 and 14 p.i. are the days of peak gameto-
cyte densities in CQ-treated and untreated infec-
tions, respectively [7]. Asexual parasites and
gametocytes per r.b.c. and r.b.c. density were
determined for each mouse on both feed days;
parasite densities were calculated from the pro-
duct of these variables. Approximately 25 mos-
quitoes per mouse were dissected 8±9 days after
Fig. 1. Mean21 S.E.M. gametocyte densities in chloroquine-
feeding and the number of parasites (oocysts) on
treated and untreated infections on days 12 and 14 p.i.
All analyses were carried out using GLIM 4.
within each analysis [19, 20]. Data from individ-
Proportions of mosquitoes infected per mouse
ual mosquitoes were nested within mouse to
were analysed by logistic regression with a bino-
avoid pseudo-replication. Non-signi®cant inter-
mial error structure. Over-dispersion in the data
actions were combined with the error term to
was corrected by using a heterogeneity factor
determine signi®cance of main eects. Dierences
(HF) of 3.05, determined from the ratio of
Pearson's w 2 to the degrees of freedom in the
between days and treatments were determined by
minimal model [19]. Day p.i. of feed (12 or 14),
ANOVA, using stepwise deletion as above, with
treatment (CQ + , CQÀ) and experiment (1, 2 or
treatment, day and experiment ®tted as factors.
3) were ®tted as factors, and square-root arcsin-
Gametocyte densities were 3.8 times higher on
transformed gametocytes per r.b.c. (gametocytae-
day 12 p.i. than day 14 p.i. in CQ-treated infec-
mia) ®tted as a covariate. The minimal model
tions, and 4.4 times higher on day 14 p.i. than
was determined by stepwise deletion, starting
day 12 p.i. in untreated infections (Fig. 1, day by
with the highest order interactions, with non-sig-
treatment interaction: F1,29 = 5.03, P < 0.05). In
ni®cant terms discarded [19]. The explanatory
the full logistic regression model, gametocytaemia
power of remaining terms was determined by del-
explained 46% of the deviance associated with
etion from the minimal model. Additional covari-
ates [log10-transformed gametocyte densities,
asexual densities and r.b.c. densities, and square-
root arcsin-transformed asexuals per r.b.c.
(asexual parasitaemia)] were substituted with
gametocytaemia in the minimal model to deter-
mine if they explained a signi®cant amount of
Further analyses were carried out in the same
way within days (HF = 3, 3.6 for days 12 and
14 p.i., respectively) and treatments (HF = 1.8,
4.1 for CQ+ and CQÀ, respectively), using only
covariates that were signi®cant in the previous
model. Analyses of oocyst burdens were also car-
ried out within days and treatments using a nega-
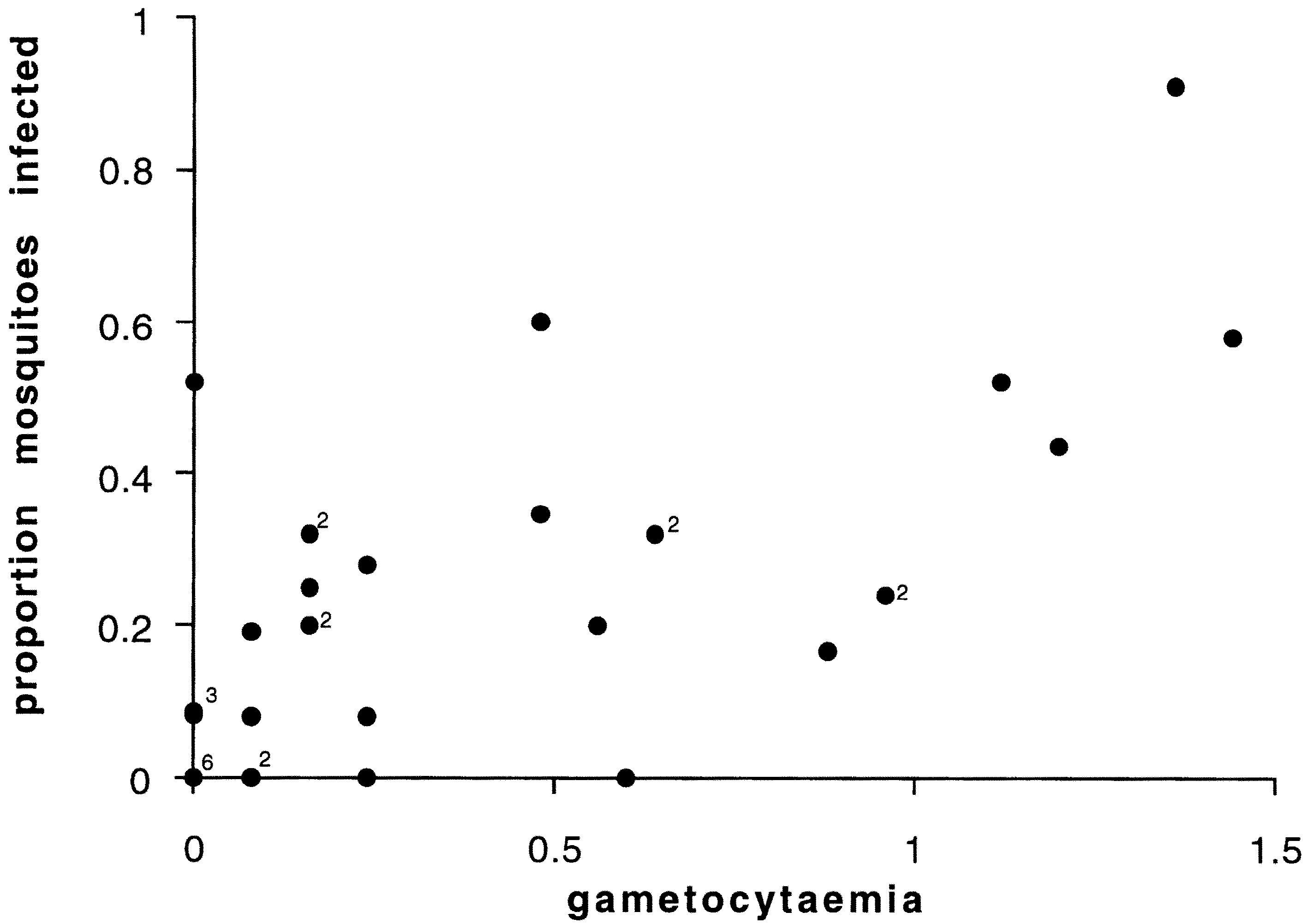
Fig. 2. Relationship between proportion of mosquitoes
infected and gametocytaemia (gametocytes per 1000 r.b.c.) for
tive binomial error structure, with the over-
all data. Small numbers in body of ®gure show multiple data
dispersion parameter, k, estimated separately
A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
proportions of mosquitoes infected (Fig. 2,
Because of the signi®cant treatment by day in-
w 21 = 25.12, P < 0.001), which was signi®cantly
teraction in the full model, data were analysed
greater than the deviance explained by gameto-
separately within days and treatments. When the
cyte density (dierence between gametocytaemia
data from day 12 p.i. alone were analysed, infec-
and gametocyte density: w 21 = 4.87, P < 0.05).
tivity per gametocyte was 3.2 times greater (at
The relationship between (untransformed) game-
the mean gametocytaemia) in CQ-treated than
untreated infections (Fig. 3a, main eect of treat-
infected was approximately linear (Fig. 2); allow-
ing the best ®t regression to curve, by the ad-
1 = 4.3, P < 0.05). Mean oocyst burdens
(per gametocyte) were 3.1 times greater (at the
dition of the quadratic function, did not explain
mean gametocytaemia) in CQ-treated compared
signi®cantly more deviance (P > 0.1).
To determine the eect of CQ on per gameto-
with untreated infections (main eect of treat-
cyte infectivity, gametocytaemia had to be con-
ment: F1,10 = 12.12, P < 0.01). The rates of
trolled for. When this was done, the eect of
increase of both proportion of mosquitoes
CQ-treatment on proportions of mosquitoes
infected and mean oocyst burdens with gameto-
infected diered on days 12 and 14 p.i. (treat-
cytaemia did not dier between CQ-treated and
untreated infections (treatment by gametocytae-
No main eects of the factors (day, experiment
mia interaction: P > 0.1). Neither proportions of
or treatment), covariates (gametocyte, asexual
mosquitoes infected or mean oocyst burdens (per
and r.b.c. densities, and asexual parasitaemia) or
gametocyte) on day 14 p.i. diered between CQ-
their other interactions explained a signi®cant ad-
treated and untreated infections (Fig. 3b, P > 0.1
ditional amount of deviance (P > 0.05, in all
for main eects of treatment, and treatment by
Fig. 3. Relationship between logit-transformed proportions of mosquitoes infected and square-root arcsin-transformed gametocy-
taemia. Lines show OLS regressions from statistical models. Labels in bold type correspond to solid symbols. Small numbers in
body of ®gures represent multiple data points. (a) Data from day 12 p.i. (b) Data from day 14 p.i. (c) Data from all untreated
infections. (d) Data from all chloroquine-treated infections.
A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
treated infections because of the greatly reduced
sidered, the proportion of mosquitoes infected
peak asexual parasite densities [7].
and mean oocyst burdens (per gametocyte) were,
In untreated infections, reduced gametocyte
at the mean gametocytaemia, respectively 3.4 and
infectivity on day 12 p.i. was also associated with
2.9 times greater on day 14 p.i. than day 12 p.i.
low gametocyte densities. There was a similar
(Fig. 3c, main eects of day: w 21 = 7.94, P < 0.01,
pattern on day 14 p.i. in CQ-treated infections.
F1,12 = 10.89, P < 0.01, respectively; day by
The infectivity of individual gametocytes may
gametocytaemia interactions: P > 0.1, in both
therefore be a positive function of gametocyte
cases). However, within CQ-treated infections,
density, as would be the case if the probability of
gametocyte infectivity as measured by both pro-
gamete fusion in the mosquito midgut was con-
portions of mosquitoes infected and mean oocyst
siderably reduced at low gametocyte densities.
burdens, did not dier between days 12 and 14
There is, however, no evidence that per gameto-
p.i. (Fig. 3d, P > 0.1 for both day and day by
cyte infectiousness increased non-linearly at high
gametocytaemia interaction, in both cases).
The results imply that if CQ does increase the
infectiousness in this study, but did CQ enhance
infectiousness of P. chabaudi gametocytes, the
per gametocyte infectiousness? On day 12 p.i.,
eect is lost by 6±7 days post-treatment. This is
gametocytes in CQ-treated mice were more infec-
before the vast majority of gametocytes were pro-
tious than those in untreated infections, both in
terms of proportion of mosquitoes infected and
infections [7]. The plasma half-life of CQ in ma-
mean oocyst burdens. Two days later, any eects
laria-infected mice is approximately 7 h [24], and
of CQ on gametocyte infectivity were no longer
thus would be at very low concentrations after
detectable. These data can be interpreted in two
6±7 days. The current results are therefore incon-
ways. First, CQ enhanced infectivity on day
sistent with the suggestion that long-term metab-
12 p.i., but the eect was lost by day 14 p.i.
olites of CQ are responsible for the reported
Second, infectivity was suppressed on day 12 p.i.
infectivity-enhancing eects of CQ [1, 17].
in untreated infections, but not on day 14 p.i.
Ultimately, our data are consistent with most
That infectivity was greater on day 14 p.i. than
previously published data: there is no strong evi-
day 12 p.i. in untreated infections, but there was
dence to suggest that CQ has long-term infectiv-
no dierence in infectivity between days in CQ-
ity-enhancing eects on gametocytes.
treated infections, leads to acceptance of the lat-
ter hypothesis. Thus, any eect of CQ on infec-
demonstrated convincingly only in the short term
(12 h post-treatment), in controlled experiments
The reduction in infectivity in untreated infec-
using rodent malaria parasites in vivo [12±14]. A
tions relative to CQ-treated infections on day 12
plausible explanation for these data is an
p.i. is consistent with the infection dynamics. In
immuno-suppressive eect of CQ. Chloroquine is
untreated infections a phenomenon called ``crisis''
occurs. This is a rapid reduction in parasite num-
presentation [25], and leukocyte function in
bers associated with low r.b.c. densities and
general [26]. Such mechanisms could conceivably
strong immune activity [21], during which game-
reduce anti-gametocyte immune activity, resulting
tocyte infectivity is suppressed [22] (and see
in increased gametocyte infectivity in the short
Carter et al. [23] for a related phenomenon in the
term. Infectivity-enhancing eects would be
human malaria parasite, P. vivax). Crisis
expected to last only as long as CQ (or the major
occurred between days 8 and 10 p.i. in untreated
antimalarial metabolite, desethyl-CQ) was at suf-
infections in these experiments, and it is entirely
®cient concentrations to have an immuno-
plausible that gametocyte infectivity-suppressing
suppressive eects. Infectivity enhancement
``crisis factors'' are still present by day 12 p.i.,
would be expected to increase with CQ dose, up
but not day 14 p.i. Crisis did not occur in CQ-
to a point where growing gametocytes were being
A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
destroyedÐmature gametocytes are unaected by
[7] Buckling AGJ, Taylor LH, Carlton JM-R, Read AF.
CQ [2]. This prediction is supported by inter-
Adaptive changes in Plasmodium transmission strategies
mediate doses of CQ having the greatest infectiv-
following chloroquine chemotherapy. Proc R Soc Lond
ity-enhancing eect [12]. Although a short-term
[8] Carter R, Graves PM. Gametocytes. In: Wernsdorfer
immuno-suppressive eect of CQ could be of po-
WH, McGregor I, editors. Malaria. Principles and prac-
tential importance to malaria epidemiology and
tice of malariology. Edinburgh: Churchill Livingstone,
the spread of drug-resistant genotypes, longer-
term eects, if they occur, would be of great
[9] Taylor LH, Read AF. Why so few transmission stages?
signi®cance. We ®nd no evidence of such long-
Reproductive restraint by malaria parasites. Parasitol
term eects in P. chabaudi. Further work on P.
[10] Chutmongkonkul M, Maier WA, Seitz HM. Plasmodium
falciparum is required to resolve this important
falciparum: eect of chloroquine, halofantrine and pyri-
methamine on the infectivity of gametocytes for
Anopheles stephensi mosquitoes. Ann Trop Med Parasitol
[11] Rosario VE, Vaughn JA, Murphy M, Harrod V,
Coleman R. Eect of chloroquine on the sporogonic
cycle of Plasmodium falciparum and Plasmodium ber-
ghei in Anopheline mosquitoes. Acta Leid 1988;57:53±
Research Fellowship to A.R. A.B. was supported
[12] Ramkaran AE, Peters W. Infectivity of chloroquine re-
by an MRC studentship. We are grateful to
sistant Plasmodium berghei to Anopheles stephensi
David Walliker for the use of his insectaries.
enhanced by chloroquine. Nature 1969;223:635±6.
Thanks to two anonymous referees for comments
[13] Hamidi AN. The eect of chloroquine on the infectivity
of rodent malaria parasites to Anopheles stephensi. Iran J
[14] Ichimori K, Curtis CF, Targett GAT. The eects of
chloroquine on the infectivity of chloroquine-sensitive
and -resistant populations of Plasmodium yoelii nigerien-
sis to mosquitoes. Parasitology 1990;100:377±81.
[15] Ponnudurai T, Lensen AH, Van Gemert GJ, Bensink
[1] Butcher GA. Antimalarial drugs and the mosquito
MP, Bolmer M, Meuwissen JH. Infectivity of cultured
transmission of Plasmodium. Int J Parasitol 1997;27:975±
Plasmodium falciparum gametocytes to mosquitoes.
[2] Smalley ME, Sinden RE. Plasmodium falciparum gameto-
[16] Wilkinson RN, Noeypatimanondh S, Gould DJ.
cytes: their longevity and infectivity. Parasitology
Infectivity of falciparum malaria patients for Anopheline
mosquitoes before and after chloroquine treatment.
[3] Klein TA, Tada MS, Lima JB, Katsuragawa TH.
Trans R Soc Trop Med Hyg 1976;70:306±7.
Infection of Anopheles darlingi fed on patients infected
[17] Hogh B, Gamage-Mendis A, Butcher GA et al. The
with Plasmodium vivax before and during treatment with
diering impact of chloroquine and pyrimethamine/sul-
chloroquine in Costa Marques, Rondonia, Brazil. Am J
fadoxine upon the infectivity of malaria species to the
mosquito vector. Am J Trop Med Hyg 1998;58:176±
[4] Bishop A. The eect of sulphadiazine, proguanil and 2:4-
diamino-6:7-diisopropylpteridine upon gametocyte pro-
[18] Tchuinkam T, Mulder B, Dechering K et al.
duction in Plasmodium gallinaceum. Parasitology
with Plasmodium falciparum of naturally infected
[5] Lines JD, Wilkes TJ, Lyimo EO. Human malaria infec-
gametocyte carriers in Cameroon: factors in¯uencing the
tiousness measured by age-speci®c sporozoite rates in
[19] Crawley M. GLIM for ecologists. Oxford: Blackwell
[6] Handunnetti SM, Gunewardena DM, Pathirana PPSL,
Ekanayake K, Weerasinghe S, Mendis KN. Features of
[20] Wilson K, Grenfell BT. Generalized Linear Modelling
recrudescent chloroquine-resistant Plasmodium falciparum
for parasitologists. Parasitol Today 1997;13:33±8.
infections confer a survival advantage on parasites and
[21] Jarra W, Brown KN. Protective immunity to malaria:
have implications for disease control. Trans R Soc Trop
studies with cloned lines of rodent malaria in CBA/Ca
mice. IV. The speci®city of mechanisms resulting in crisis
A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
and resolution of the primary acute phase parasitaemia
[24] Cambie G, Verdier F, Gaudebout C, Clavier F,
of Plasmodium chabaudi chabaudi and P. yoelii yoelii.
Ginsburg H. The pharmacokinetics of chloroquine in
healthy and Plasmodium chabaudi-infected mice: impli-
[22] Wery M. Studies on the sporogony of rodent malaria
cations for chronotherapy. Parasite 1994;1:219±26.
parasites. Ann Soc Belge Med Trop 1968;48:1±138.
[25] Fox RI, Kang H-I. Mechanism of action of antimalarial
[23] Carter R, Wijesekera SK, Karunaweera ND, Mendis
drugs: inhibition of antigen processing and presentation.
KN. Mediators and mechanisms associated with parox-
ysm in Plasmodium vivax malaria. J Pharm Pharmacol
[26] Targett GAT. Drug use and the immune response.
First Name:__________________________ M.I. _____ Last Name:____________________________ Male / Female D.O.B:_________________ Employer: _____________________________ MARRIED / SINGLE / OTHER Home Phone: _______________________ Cell: _____________________ Work: ________________________ Address: ____________________________City/State/Zip Code: _____________________________________________ Prim
SIVA SIVANI INSTITUTE OF MANAGEMENT From Director’s Desk News Brief 22 -07-2010 __________________________________________________________________ MARKETING 1. Toyota launches diesel variant of Corolla Altis Gearing up for December launch of its small car, Etios. Toyota Kirloskar Motors (TKM) on Wednesday launched the diesel version of its premiumsedan, the Corolla Altis. Priced at

 International Journal for Parasitology 29 (1999) 619±625
The eect of chloroquine treatment on the infectivity of
Institute of Cell, Animal and Population Biology, Division of Biological Sciences, University of Edinburgh, Edinburgh EH9 3JT, UK
Received 19 October 1998; received in revised form 3 December 1998; accepted 3 December 1998
The antimalarial drug chloroquine has been reported to increase the infectivity of the forms of blood-stage
malaria parasites (gametocytes) that are capable of infecting mosquito vectors. This eect has been demonstrated
convincingly in the short term (12 h post treatment), although several authors have suggested infectivity
enhancement a week or more after treatment. We carried out experiments to investigate the eects of chloroquine
on the longer-term infectivity of gametocytes of the rodent malaria parasite, Plasmodium chabaudi, to Anopheles
stephensi mosquitoes. Gametocytes of chloroquine-treated infections were signi®cantly more infectious than
untreated infections 6 and 7 days post-treatment, although not on days 8 and 9. However, this eect was most likely
the result of a reduction in infectivity in untreated infections, caused by immune activity which was not so
pronounced in chloroquine-treated infections. Gametocytaemia (gametocytes per r.b.c.) showed a strong positive
and linear relationship with infectivity. Infectivity was not in¯uenced by either asexual parasitaemia, asexual density
or anaemia. Parsimonious interpretations of the eect of chloroquine on gametocyte infectivity are discussed.
International Journal for Parasitology 29 (1999) 619±625
The eect of chloroquine treatment on the infectivity of
Institute of Cell, Animal and Population Biology, Division of Biological Sciences, University of Edinburgh, Edinburgh EH9 3JT, UK
Received 19 October 1998; received in revised form 3 December 1998; accepted 3 December 1998
The antimalarial drug chloroquine has been reported to increase the infectivity of the forms of blood-stage
malaria parasites (gametocytes) that are capable of infecting mosquito vectors. This eect has been demonstrated
convincingly in the short term (12 h post treatment), although several authors have suggested infectivity
enhancement a week or more after treatment. We carried out experiments to investigate the eects of chloroquine
on the longer-term infectivity of gametocytes of the rodent malaria parasite, Plasmodium chabaudi, to Anopheles
stephensi mosquitoes. Gametocytes of chloroquine-treated infections were signi®cantly more infectious than
untreated infections 6 and 7 days post-treatment, although not on days 8 and 9. However, this eect was most likely
the result of a reduction in infectivity in untreated infections, caused by immune activity which was not so
pronounced in chloroquine-treated infections. Gametocytaemia (gametocytes per r.b.c.) showed a strong positive
and linear relationship with infectivity. Infectivity was not in¯uenced by either asexual parasitaemia, asexual density
or anaemia. Parsimonious interpretations of the eect of chloroquine on gametocyte infectivity are discussed.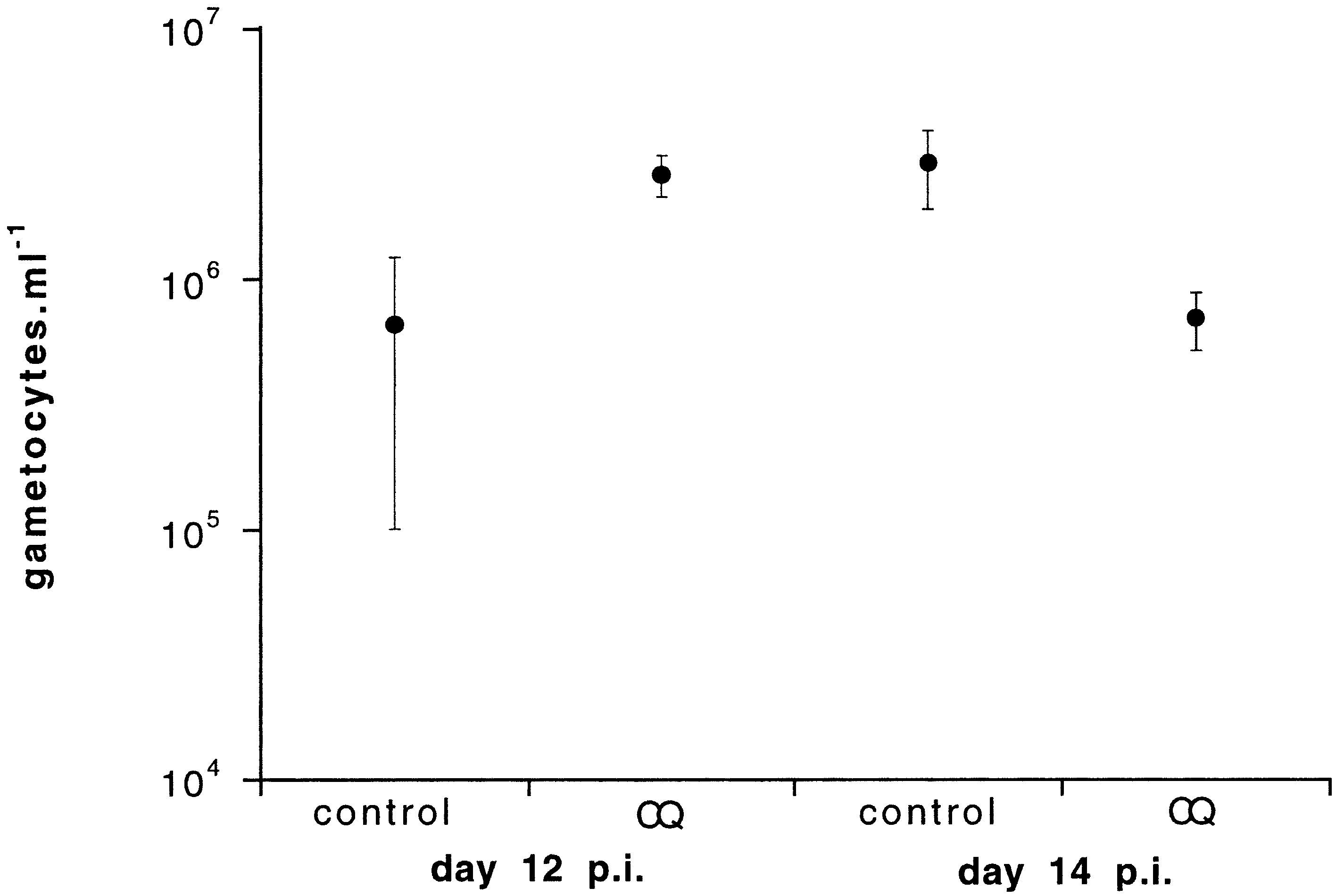
 A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
mice were picked randomly and fed to H40 4±5-
day-old Anopheles stephensi on day 12 p.i. (except
in the control group in experiment 1, where two
animals out of ®ve were fed to mosquitoes). The
other mice were fed to mosquitoes on day 14 p.i.
A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
mice were picked randomly and fed to H40 4±5-
day-old Anopheles stephensi on day 12 p.i. (except
in the control group in experiment 1, where two
animals out of ®ve were fed to mosquitoes). The
other mice were fed to mosquitoes on day 14 p.i.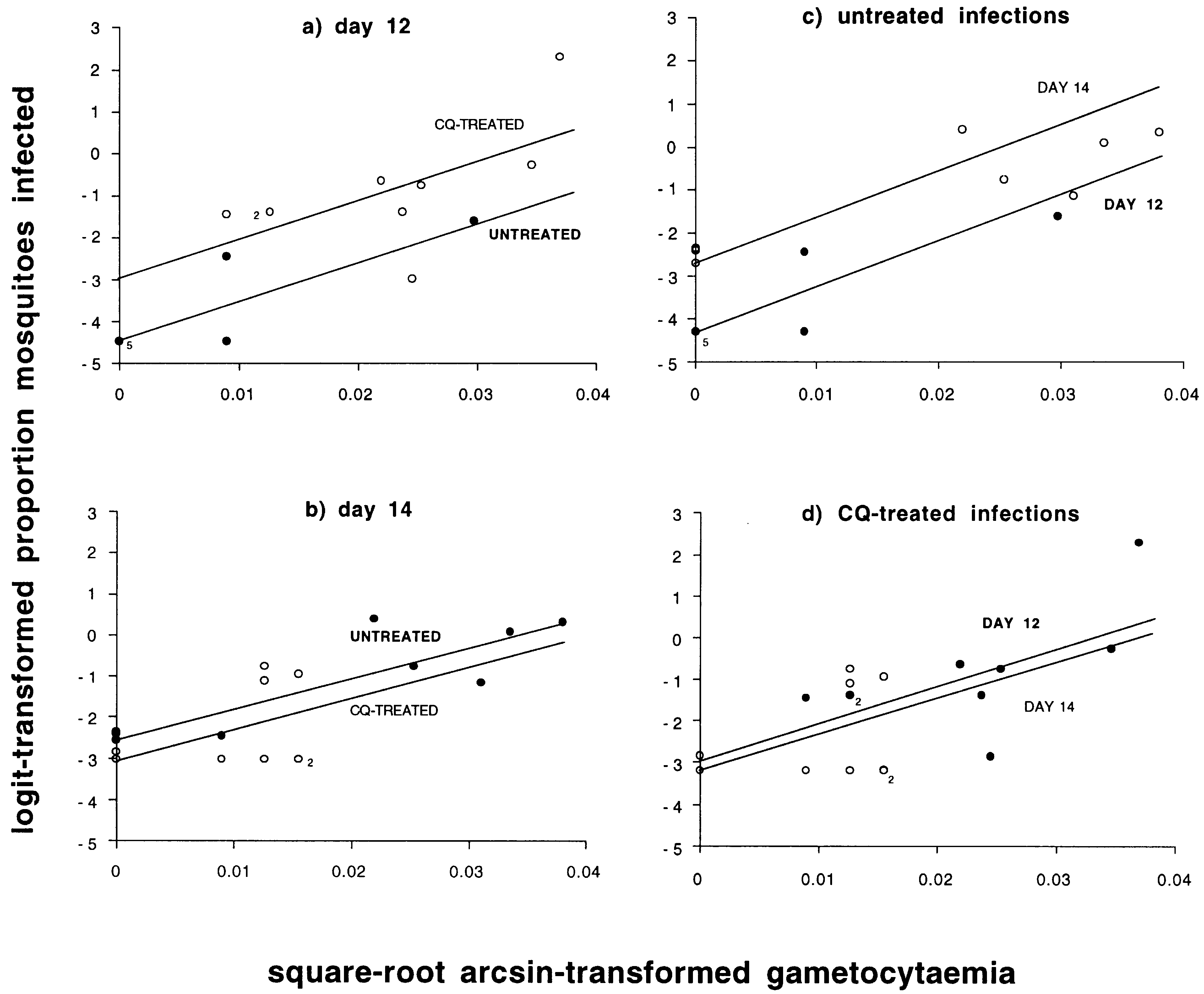 A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
proportions of mosquitoes infected (Fig. 2,
Because of the signi®cant treatment by day in-
w 21 = 25.12, P < 0.001), which was signi®cantly
teraction in the full model, data were analysed
greater than the deviance explained by gameto-
separately within days and treatments. When the
cyte density (dierence between gametocytaemia
data from day 12 p.i. alone were analysed, infec-
and gametocyte density: w 21 = 4.87, P < 0.05).
A.G.J. Buckling, A.F. Read / International Journal for Parasitology 29 (1999) 619±625
proportions of mosquitoes infected (Fig. 2,
Because of the signi®cant treatment by day in-
w 21 = 25.12, P < 0.001), which was signi®cantly
teraction in the full model, data were analysed
greater than the deviance explained by gameto-
separately within days and treatments. When the
cyte density (dierence between gametocytaemia
data from day 12 p.i. alone were analysed, infec-
and gametocyte density: w 21 = 4.87, P < 0.05).