Soy Diets Containing Varying Amounts of Genistein Stimulate Growth of Estrogen-dependent (MCF-7) Tumors in a Dose-dependent Manner
Clinton D. Allred, Kimberly F. Allred, Young H. Ju, et al. Updated Version
Access the most recent version of this article at:
Cited Articles
This article cites 32 articles, 22 of which you can access for free at:
Citing Articles
This article has been cited by 44 HighWire-hosted articles. Access the articles at:
E-mail alerts Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Subscriptions Permissions
To request permission to re-use all or part of this article, contact the AACR PublicationsDepartment at
[CANCER RESEARCH 61, 5045–5050, July 1, 2001]
Soy Diets Containing Varying Amounts of Genistein Stimulate Growth of Estrogen- dependent (MCF-7) Tumors in a Dose-dependent Manner1 Clinton D. Allred,2 Kimberly F. Allred,2 Young H. Ju, Suzanne M. Virant, and William G. Helferich3 Department of Food Science and Human Nutrition [C. D. A., K. F. A., Y. H. J., S. M. V., W. G. H.], and Division of Nutritional Sciences [C. D. A., W. G. H.], University of Illinois,Urbana, Illinois 61801ABSTRACT
various forms, including supplement capsules, which contain mixturesof isoflavones derived from soy. Isoflavones are also present in
We have demonstrated that the isoflavone, genistein, stimulates growth
bioactive concentrations in food products such as soy protein isolates,
of estrogen-dependent human breast cancer (MCF-7) cells in vivo (C. Y.
which can contain varying isoflavone content. It is generally accepted
Hsieh et al., Cancer Res., 58: 3833–3838, 1998). The isoflavones are a group of phytoestrogens that are present in high concentrations in soy.
that consuming the phytochemical components of soy, particularly the
Whether consumption of genistein from soy protein will have similar
isoflavones, in pure form, as in supplements, may pose some health
effects on estrogen-dependent tumor growth as pure genistein has not
concerns but that consumption of more whole foods containing these
been investigated in the athymic mouse tumor implant model. Depending
are natural and, as such, safe. It is important to note that the concen-
on processing, soy protein isolates vary widely in concentrations of genis-
tration of these compounds in soy protein isolates is dependent on the
tein. We hypothesize that soy isolates containing different concentrations
method of processing, and specific processing methods can be used to
of genistein will stimulate the growth of estrogen-dependent cells in vivo in
enrich the isoflavone content in soy protein isolates. a dose-dependent manner. To test this hypothesis we conducted experi-
Clinical and preclinical laboratory animal and in vitro studies have
ments in which these soy protein isolates were fed to athymic mice
demonstrated the hormonal activity of dietary isoflavones. Our labo-
implanted s.c. with estrogen-dependent tumors. Genistein content (agly- cone equivalent) of the soy isolate diets were 15, 150, or 300 ppm. Positive
ratory has examined the estrogenic activity of genistein. In vitro, when
(with 17-estradiol pellet implant) and negative (no 17-estradiol) control
human estrogen-dependent breast cancer (MCF-7) cells were treated
groups received casein-based (isoflavone-free) diets. Tumor size was meas-
with increasing concentrations of genistein (1–10 M), the cells ex-
ured weekly. At completion of the study animals were killed and tumors
pressed increased mRNA levels of pS2, an estrogen-responsive gene,
collected for evaluation of cellular proliferation and estrogen-dependent
in a dose-dependent manner. In vivo, 25-day-old ovariectomized,
gene expression. Incorporation of bromodeoxyuridine into cellular DNA
athymic mice consuming dietary genistein for 5 days had increased
was used as an indicator of cell proliferation, and pS2 mRNA was used as
number and size of terminal end buds in the mammary gland when
an estrogen-responsive gene. Soy protein diets containing varying
compared with control animals (5). Not only do the isoflavones have
amounts of genistein increased estrogen-dependent tumor growth in a
estrogenic activity on various tissues when given in pure form but also
dose-dependent manner. Cell proliferation was greatest in tumors of animals given estrogen or dietary genistein (150 and 300 ppm). Expression
when consumed in products such as soy protein isolate. In humans,
of pS2 was increased in tumors from animals consuming dietary genistein
Petrakis et al. (6) demonstrated that consumption of soy protein
(150 and 300 ppm). Here we present new information that soy protein
isolate had stimulatory effects on the breast tissue of premenopausal
isolates containing increasing concentrations of genistein stimulate the
women. They found that duct fluid aspirates contained greater num-
growth of estrogen-dependent breast cancer cells in vivo in a dose-depen-
bers of hyperplastic epithelial cells in women consuming soy protein
dent manner.
isolate. Nipple aspirate apolipoprotein D and pS2 expression has alsobeen shown to be elevated in women consuming 60 g of soy in the
INTRODUCTION
form of ground, textured vegetable protein for as little as 2 weeks (7). These data collected from both laboratory animals and humans dem-
The Food and Drug Administration recently approved a health
onstrate the estrogenic activity of the isoflavones whether they are
claim for soy protein. The claim states “25 g of soy protein a day, as
taken as a pure compound or consumed in soy protein isolate con-
part of a diet low in saturated fat and cholesterol, may reduce the riskof heart disease” (1). As a result, the consumption of soy protein by
Americans has increased. Soy and soy-derived supplements are not
The role that isoflavones play in breast cancer is unclear. Some
being used solely to lower serum cholesterol and reduce risk of heart
reports indicate that exposure to genistein is preventative in the
disease. Soy contains a complex mixture of a variety of phytochemi-
development of breast cancer (8 –10), whereas others show that ge-
cals; our focus is on the estrogenic isoflavones referred to as phy-
nistein stimulates the growth of existing estrogen-dependent tumors
toestrogens. Isoflavones have been demonstrated to act as estrogen
(5). Studies published over the last 5 years have demonstrated that
agonists by binding to the estrogen receptor and generating estrogen-
exposure to dietary genistein before puberty reduces the number of
induced responses (2– 4). As a result, postmenopausal women may
chemically induced mammary tumors formed in female Sprague
consume soy for the estrogenic effects of these compounds to relieve
Dawley rats. The authors suggest protection against the development
menopausal symptoms. Soy products are marketed as a “natural”
of breast cancer tumors is attributable to the estrogenic effects of
alternative to hormone replacement therapy with the perception that
genistein that causes increased cellular differentiation in mammary
these phytoestrogens are without the risks associated with hormone
gland cells of prepubertal animals. A differentiated cell undergoes less
replacement therapy. Isoflavone-containing products are produced in
proliferation and therefore is less likely to progress through the cancerprocess (8 –10). If prepubertal exposure to genistein results in earlierdifferentiation of the mammary gland, then the assumption can be
Received 8/9/00; accepted 4/27/01. The costs of publication of this article were defrayed in part by the payment of page
made that earlier differentiation would be protective against chemi-
charges. This article must therefore be hereby marked advertisement in accordance with
cally induced mammary tumors. Cohen et al. (11) found that continual
18 U.S.C. Section 1734 solely to indicate this fact.
1 Supported by NIH Grant CA77355 (to W. G. H.).
feeding of soy protein isolate (containing 1.67 mg total isoflavones
2 C. D. A. and K. F. A. contributed equally to this paper.
per g isolate) postpubertally for 18 weeks after chemical induction of
3 To whom requests for reprints should be addressed, at 580 Bevier Hall, Department
mammary tumors in rats had no detectable effect on the development
of Food Science and Human Nutrition, University of Illinois, Urbana, IL 61801. Phone:(217) 244-5414; Fax: (217) 244-7877; E-mail: helferic@uiuc.edu.
of these tumors. Our research has focused on the effect dietary
SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
genistein has on the growth of existing estrogen-dependent (MCF-7)
diets. Treatment animals were fed either soy isolate-containing diets or AIN-
tumors in animals with low circulating E 4 concentrations. We have
93G diet plus genistein. Three soy protein isolates with varying concentrations
demonstrated that genistein fed at 750 ppm stimulates the growth of
of genistein were used in this study. Each isolate was added at 20% of the diet
MCF-7 cells implanted into athymic mice. Mice consuming 750 ppm
as the sole source of protein in the AIN-93G diet. Both the casein-based diet
dose of genistein have a total plasma genistein concentration of ϳ2
and the soy-based diet provided ϳ18% protein, but 2.2 g of methionine was
added to each kilogram of soy isolate diet to meet the sulfur-containing amino
M (5). This is a relevant dietary dosage because women who
acid requirements and as a result, all of the essential amino acid requirements
consume varying amounts of isoflavones from soy milk have plasma
of the mouse were met. The three soy protein isolates (Protein Technologies
genistein levels of 0.8-2.2 M (12). In vitro genistein has been shown
International, St. Louis, MO) were processed to contain varying amounts of
to stimulate the growth of MCF-7 cells at concentrations as low as 200
genistein. There were no significant differences in amino acid content or
nM (2, 5). Whereas these data demonstrate genistein can stimulate the
proximate analysis among the isolates. Additionally, the isolates were very low
growth of estrogen-dependent breast cancer tumors, it has not yet been
in fiber and as such the lignin content was negligible. The majority of the
determined if soy protein isolate, which contains a complex mixture of
isoflavones in the protein isolates were present as the aglycone form (ϳ60%).
phytochemicals, can exhibit similar effects on growth of estrogen-
The genistein contents were 0.075, 0.75, and 1.5 mg genistein (aglycone
equivalents)/g of product. This resulted in the final concentrations of genistein
The study presented here was designed to evaluate the effects of
in the three soy isolate diets to be 15, 150, and 300 mg genistein/kg diet
dietary soy protein isolates containing varying concentrations of ge-
respectively. To compare the genistein content from soy to pure genistein, itwas added to the AIN-93G diet at equal concentrations to that found in the soy
nistein: (a) on the growth of MCF-7 tumors in athymic mice; (b) on
isolate diets and provided 15, 150, and 300 ppm genistein in a casein-based
cellular proliferation of estrogen-dependent tumors in ovariectomized
diet. The eight final treatment groups were: positive controls (2 mg E pellet);
athymic mice; and (c) on expression of the estrogen-responsive gene,
negative controls; low soy isolate [LSI (15 ppm genistein)]; medium soy
pS2, in response to dietary exposure to soy protein isolates with
isolate [MSI (150 ppm genistein)]; high soy isolate [HSI (300 ppm genistein)];
varying concentrations of genistein.
low genistein [LG (AIN-93G ϩ 15 ppm genistein)]; medium genistein [MG(AIN-93G ϩ 150 ppm genistein)]; and high genistein [HG (AIN- 93G ϩ 300ppm genistein)]. MATERIALS AND METHODS E Pellet Preparation. MCF-7 cells will not produce tumors in ovariec-
tomized mice unless they are supplemented with estrogen. Therefore, 1 week
Effects of Consumption of Soy Protein Isolate and Casein-based Diets
after delivery, animals were implanted with E pellets. E pellets were made
Containing Genistein on the Growth of Estrogen-dependent Tumors
containing 2 mg of E mixed with 18 mg of cholesterol as a carrier. A 20-mg
mixture containing E and cholesterol was placed into a pellet mold and
Animals. Female athymic nude mice were purchased from Harlan Sprague
pressed into a compact pellet ϳ4.5 mm in diameter and ϳ2.5 mm in depth.
Dawley (Indianapolis, IN) and delivered at 28 days of age. Mice were ovari-ectomized at 21 days of age and allowed a week to recover before delivery. A
Pellets were then placed s.c. in the interscapular region of mice (14).
2-mg E pellet was placed under the skin of each mouse before MCF-7 cells
Tumor Implantation. MCF-7 cells were maintained in 100 mm ϫ 20 mm
were transplanted into the animal. Cells were then injected into the four flanks
plastic tissue culture plates in IMEM media (Biofluids) containing 10% fetal
on the back of each animal. Within the first 4 weeks, tumors that subsequently
bovine serum, penicillin (100 units/ml) and streptomycin (100 units/ml). E2
formed were allowed to develop to an average cross-sectional area of ϳ40
(1.0 nM) was added in the media to keep the cell line estrogen-dependent. Cells
mm2. At this point, animals were placed into eight treatment groups (seven
were incubated at 37°C in a humidified atmosphere of 5% CO . Cells were
animals/group). The treatment groups were: (a) positive control; (b) negative
grown to confluence, collected using trypsin-EDTA, and counted. Cells were
control; (c) dietary soy protein isolate providing 15 ppm genistein; (d) dietary
suspended in matrigel matrix (Becton Dickinson) before being injected (15)
soy protein isolate providing 150 ppm genistein; (e) dietary soy protein isolate
and then 40 l of the cell suspension 1.5 ϫ 105 cells were injected per site into
providing 300 ppm genistein; (f) dietary genistein in casein-based diet at 15
each of the four flanks of the athymic mice.
ppm; (g) 150 ppm; and (h) 300 ppm. The E pellet was removed from the
Tumors were measured weekly and cross-sectional area was determined
negative control animals and from all of the mice on the various dietary
using the formula [length/2 ϫ width/2 ϫ ] (5, 16). When tumors reached an
treatments. The E pellet was also removed from the positive control group,
average cross-sectional area of 40 mm2, animals were divided into treatment
but these animals were again implanted with an identical 2-mg E pellet.
groups with each group normalized for tumor number, tumor size, and animal
Negative and positive controls were given AIN 93G as a control diet. This diet
number. E pellets were removed from all of the animals, and the mice were
provided protein from casein. The remaining animals were put on one of the
then placed on the treatment diets. Positive control mice were reimplanted with
six treatment diets. Tumor area and body weight were measured weekly. At the
a fresh pellet containing 2 mg of E. We then resumed measuring tumor areas
end of the study, tumors and plasma samples were collected for tissue analysis.
We felt that weekly tumor area was the most critical outcome evaluated. Tissue/Tumor Collection. At the completion of the study, mice were
Measurements were made over a total of 29 weeks after removal of E pellets.
killed by cervical dislocation, and tumors were harvested. Tumors from each
However, at the conclusion of the study, tumors from the negative control, low
mouse were fixed in 10% formalin and embedded for immunohistochemical
soy isolate, and low genistein groups were small, making it impossible to
staining. Additional tumors from each mouse were immediately frozen in
collect representative samples for mRNA and cell proliferation data. There-
liquid nitrogen for subsequent RNA isolation and analysis.
fore, to obtain these measurements for the negative control group, a second setof negative control animals were used. These animals were killed 11 weeks
Analysis of Estrogen-responsive pS2 mRNA
after removal of the E pellet when their tumors were at an average cross-
sectional area of 25 mm2. We feel this was the most accurate method to obtain
RNA Preparation. The mRNA was isolated using procedures routinely
tissues from the negative control group that were regressing but still large
used in our laboratory (17). Briefly, frozen tumors (Յ200 mg) from liquid
enough to obtain data for mRNA and immunohistochemical analysis.
nitrogen were smashed and the coarse tumor powder was transferred into
Diet Formulation. AIN-93G semipurified diet was selected as a base diet
TRIZOL (Life Technologies, Inc., Grand Island, NY) in a 15-ml tube and was
for control animals as it has been established as meeting all of the nutritional
homogenized using a Polytron-Aggregate (Luzern, Switzerland). Chloroform
requirements of mice (13). Soy oil was removed from all of the diets and corn
was added into a homogenized tumor sample, shaken vigorously, and then
oil added to eliminate any additional components of soy being added to the
incubated for 10 min at 24°C. The reaction tube was centrifuged at 12,000 ϫ gfor 15 min at 4°C. The upper portion was removed and transferred into a freshtube. An equal volume of isopropyl alcohol was added, shaken, and incubated
4 The abbreviations used are: E , 17-estradiol; BrdUrd, 5-bromo-2Ј-deoxyuridine;
for 10 min at 24°C. The mixture was centrifuged at 12,000 ϫ g for 10 min at
LSI, low soy isolate; MSI, medium soy isolate; HSI, high soy isolate; LG, low genistein;
4°C. The RNA pellet was washed with ice-cold 75% ethanol and centrifuged
MG, medium genistein; HG, high genistein; AIN 93G, American Institute of Nutrition 93growth diet.
at 7,500 ϫ g for 5 min at 4°C. The RNA pellet was air-dried then dissolved
SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
with RNase-free dH O. RNA was stored at Ϫ80°C. RNA concentration was
ϭ 40 g of single-stranded RNA/ml). Northern Blot Analysis. Expression of pS2 was used as a biomarker of
estrogenic activity (18). For the detection of pS2 expression, 10 g of RNAwere separated on 1.2% formaldehyde denaturing agarose gels and transferredto a Magnacharge, Nylon, Transfer membrane (Osmonics, Westboro, MA). The RNA was UV cross-linked onto the membrane. The membrane wasprehybridized in a formamide prehybridization solution containing denaturedsalmon sperm DNA for 3 h at 42°C. After 3-h prehybridization, the DNAprobe was labeled using Random Primers DNA Labeling System (Life Tech-nologies, Inc.). For the estrogen-responsive pS2 gene, a 25 ng of pS2 cDNA,or for the control, a 25 ng of glyceraldehyde-3-phosphate dehydrogenasecDNA probe was labeled with 50 Ci ␣-[ 32P]dCTP. The membrane wasincubated with 32P-labeled probe overnight at 42°C. The membrane waswashed twice with 5 ml of 0.2% SSC/0.1% SDS at 24°C for 5 min each, threetimes with 5 ml of 0.1% SSC/0.1% SDS at 68°C for 15 min each, and rinsedwith 2% SSC. The membrane was exposed to X-ray film for 6 h, andhybridizing RNA molecules were detected by performing autoradiography. Film was analyzed using Collage (version 4.0) software (Image DynamicsCorp.) with Foto Analyst (Futodyne). Tumor Cell Proliferation Immunohistochemistry BrdUrd Analysis. BrdUrd incorporation into cellular DNA was used as an
indicator of cells that were actively proliferating (19). Four h before killing theanimals each mouse was injected i.p. with 50 mg BrdUrd/kg body weight. Tumors were excised, skin and fat removed, and processed for tissue staining. Prepared sections were then stained for the presence of BrdUrd using amodified immunohistochemistry protocol (20). Briefly, slides were deparaf-finized and hydrated by immersing in xylene twice for 12 min and immersingin a series of alcohol/water solutions for 5 min each. To block endogenousperoxidase, slides were immersed in 0.3% H O for 20 min then washed with
distilled water. Slides were then microwaved in a Pyrex dish in 18 mM citratebuffer at pH 6.0 for 20 min and cooled. Then slides were washed in PBS (pH7.1–7.4) for 5 min, and tissue sections were rimmed with wax. 50-l anti-BrdUrd primary antibody (Amersham) was added to slides and incubated for1 h at room temperature in a humidity chamber. Slides were washed in PBSand 50 l of diluted secondary antibody (Sigma Chemical Co.) was added toslides and incubated for 30 min at room temperature. Slides were then washed
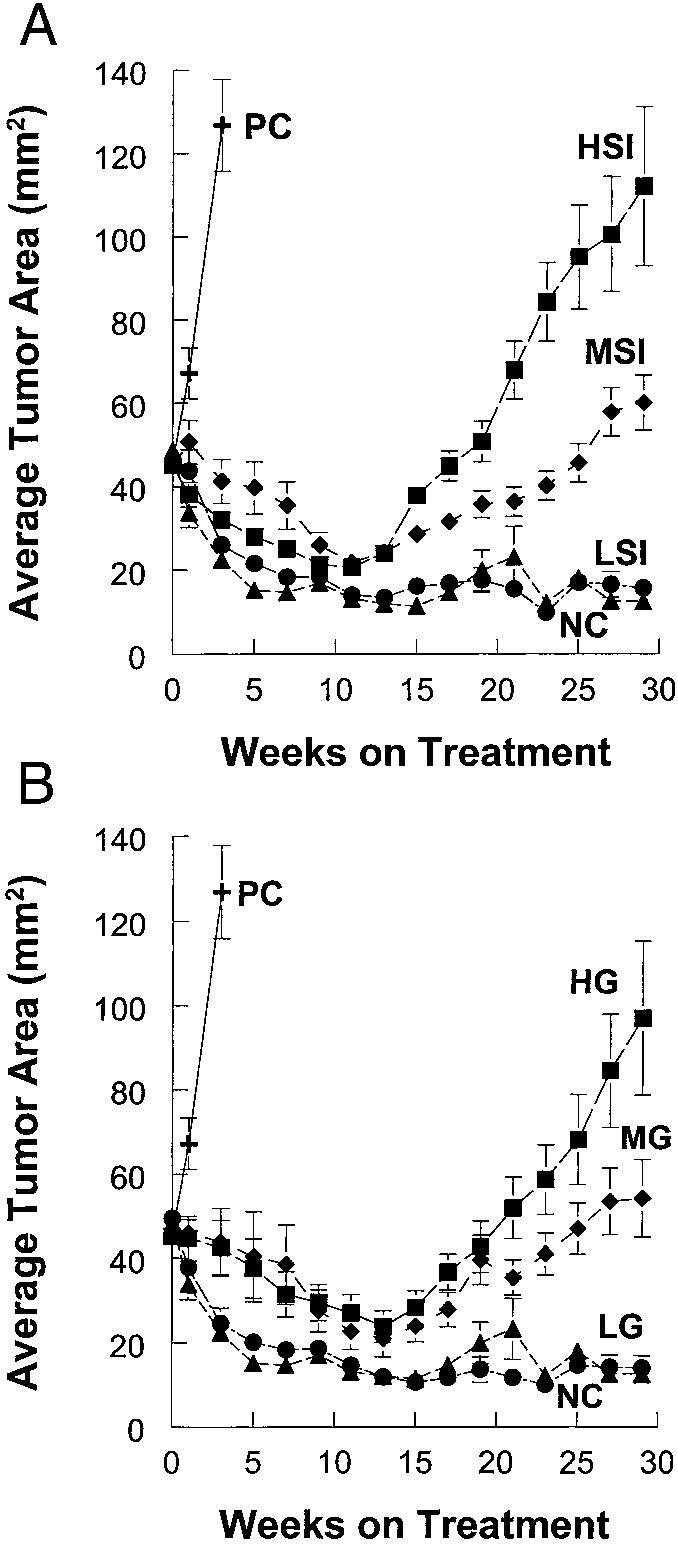
Fig. 1. A and B, effects of soy protein isolates and genistein on MCF-7 tumor growth
in athymic mice. Female ovariectomized athymic mice were implanted with a 2-mg E
in PBS. One drop of 3,3Ј-diaminobenzidine ϩ Ni enhancer solution, freshly
pellet. The animals were then injected with 1.5 ϫ 105 MCF-7 cells/site in four locations.
prepared, was added to each slide with a Pasteur pipette. Slides were then
Subsequently, tumors developed and were allowed to grow to an average cross-sectional
washed in water twice, in PBS, and counterstained with 20% hematoxylin for
area of 40 mm2. At this time, E pellets were removed from all of the mice, and they were
1 min. The slides were then dehydrated by placing them in 80% alcohol for 5
assigned to one of eight treatment groups: positive controls that were reimplanted with anew 2-mg E pellet (8 mice; n ϭ 32 tumors), negative controls that were fed AIN 93G
min, 95% ethanol for 5 min, and 100% ethanol for 5 min followed by xylene
rodent diet alone (6 mice; n ϭ 24 tumors), LSI containing 15 ppm genistein (7 mice;
four times for 5 min each. Slides were then coverslipped and analyzed by light
n ϭ 27 tumors), MSI containing 150 ppm genistein (8 mice; n ϭ 31 tumors), HSI
microscope. Both positive and background stained cells were counted in a
containing 300 ppm genistein (6 mice; n ϭ 23 tumors), LG AIN 93G ϩ 15 ppm genistein
given area of tissue. The data were then presented as percentage of cells
(7 mice; n ϭ 27 tumors), MG AIN 93G ϩ 150 ppm genistein (6 mice; n ϭ 23 tumors),and HG AIN 93G ϩ 300 ppm genistein (7 mice; n ϭ 27 tumors). A, the effects of soy
protein isolate on the growth of MCF-7 tumors. B, the effect of consumption genistein in
Statistical Analysis. Tumor area data were analyzed using one-way or
AIN 93G on the growth of MCF-7 tumors. In soy-containing diets, the soy protein isolates
repeated-measures ANOVA according to the characteristics of the data set
were the sole source of protein whereas casein was the protein source in AIN93G diets
using the SAS program. pS2 gene expression data were analyzed using
supplemented with genistein. Both soy- and casein-based diets were formulated to meetall of the nutritional requirements of the mice. The day animals were started on experi-
one-way treatment ANOVA. If the overall treatment F-ratio was significant
mental diets and were designated as measurement 0. Tumors were then measured weekly.
(P Ͻ 0.05), the differences between treatment means were tested with Fisher’s
Data are expressed as average cross-sectional tumor area for all of the tumors in each
and LG groups were similar to the negative control group resulting in
Effect of Soy Isolates and Genistein on MCF-7 Tumor Growth
average tumor areas of 16 mm2 and 14 mm2, respectively. The
in Athymic Mice. By week 3 after retreatment with a new 2-mg E
average tumor area in the MSI and MG groups were significantly
pellet, the average cross sectional area of the tumors in the positive
higher than the negative control group with final cross sectional areas
control group was 126 mm2 (Fig. 1, A and B). At this point, these mice
of 60 mm2 and 54 mm2 (P Ͻ 0.01) respectively. The MSI and MG
groups were not significantly different from one another. The tumors
negative control tumors regressed to an average area of 12 mm2. They
from both the HSI and HG groups were significantly larger than the
then maintained this size until termination of the study. Soy protein
negative control and other dietary treatment groups (P Ͻ 0.01). The
isolates stimulated growth of MCF-7 tumors in a dose-dependent
HSI average area was 112 mm2 whereas the HG group had a final
fashion (Fig. 1A). Also, casein-based diets containing equivalent
average area of 97 mm2. These (HSI and HG) were not significantly
levels of genistein stimulated tumor growth in a dose-dependent
different from one another and were similar to that obtained from the
manner (Fig. 1B). By week 29 after E pellet removal, both the LSI
positive control group 3 weeks after retreating them with E pellets
SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
(Fig. 2). Body weight was monitored weekly, and no significantdifference was observed among the treated and control groups (datanot shown). These data indicate that dietary soy isolates containingincreasing concentrations of genistein and casein-based diets contain-ing equalized concentrations of genistein act in a dose-dependentmanner to stimulate growth of human estrogen-dependent breastcancer cells transplanted into athymic mice. Also, it is important tonote that there were no significant differences in growth of tumors inthe negative control, low soy isolate, and low genistein groups. Thesedata suggest that there is a threshold level of dietary genistein belowwhich no increase in estrogen-dependent tumor growth is observed. Effect of Soy Isolates and Genistein on Estrogen-responsive pS2 mRNA Expression in MCF-7 Tumors. To evaluate the ability of soy isolates and genistein in the diet to enhance the expression of an
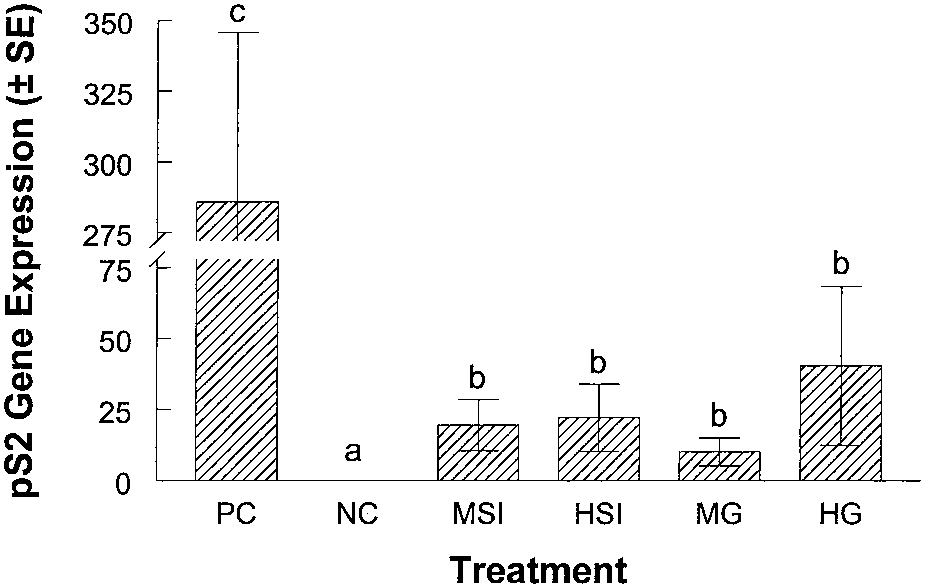
Fig. 3. Effect of soy and genistein treatments on pS2 gene expression in MCF-7
estrogen-responsive gene, pS2, we conducted Northern blot analysis
tumors. At the end of the study, tumors were collected for analysis. For the detection of
using mRNA isolated from tumors excised from animals in each
pS2 expression, mRNA was isolated from each tumor. Northern blot analysis was used toquantify the amount of mRNA produced as a result of the treatments. Three tumors from
treatment group. At the conclusion of the study, tumors from the
each treatment group were used for pS2 analysis. pS2 expression is presented as the
negative control, low soy isolate, and low genistein groups were too
relative pS2 mRNA level. Glyceraldehyde-3-phosphate dehydrogenase was used as astandard. Bars, SE.
small to obtain sufficient RNA for Northern blot analysis. Negativecontrol tumors were collected from a separate set of tumor-bearingathymic mice in which the tumors were regressed for 11 weeks after
ing and nonproliferating cells in a given field of view were counted,
E pellets were removed. This allowed us tumors that were regressing
and final values were expressed as percentage of proliferating cells.
(attributable to low E ) but were large enough for RNA isolation.
The percentage of proliferating cells value for the negative control
Expression of pS2 was not detectable in the RNA isolated from
group was 1.4%. Both the MSI and MG groups had a significantly
negative control animals. Expression of pS2 was very low in the
higher percentage of cellular proliferation when compared with the
isoflavone-treated animals, ϳone-tenth that of the E -treated mice.
negative control group with values of 9.4% and 8.2% (P Ͻ 0.001).
However, pS2 expression was significantly (P Ͻ 0.05) higher in
The animals consuming the high soy isolate with the highest concen-
animals consuming the medium and high soy protein isolates when
tration of genistein and the animals consuming the highest concen-
compared with the negative control group. Levels of pS2 expression
tration of genistein in control diet had proliferation of 11.9% and
in tumors from animals that were consuming medium and high
11.8% respectively. These values were significantly higher than all of
concentrations of genistein in the AIN93G diet were also significantly
the other groups (P Ͻ 0.05; Fig. 4).
(P Ͻ 0.05) higher than the negative control group (Fig. 3). However,there was no significant difference between the MSI and HSI groups
DISCUSSION Cellular Proliferation in MCF-7 Tumors Excised from Animals
The purpose of this study was to determine the influence of dietary
Consuming Soy Protein Isolates and Genistein. Cellular incorpo-
soy protein isolates containing increasing concentrations of genistein
ration of BrdUrd was used as an indicator of cellular proliferation in
on the growth of estrogen-dependent human breast cancer cells trans-
MCF-7 tumors. Cells that stained positive after immunohistochemical
planted into athymic mice. The results presented here demonstrate that
analysis were considered actively proliferating cells. Both proliferat-
soy protein isolates containing varying concentrations of genistein canstimulate growth of estrogen-dependent tumors similar to that seenwith pure dietary genistein. Soy protein isolates stimulated the growthof MCF-7 tumors in a dose-dependent manner as the concentration ofgenistein increased in the isolates. Tumor growth was significantlyincreased in animals consuming genistein at concentrations of 150 and300 ppm in both the casein- and soy-based diets. Dietary concentra-tions in this study are lower than previous studies in which MCF-7tumor growth was observed in athymic mice consuming dietarygenistein at 750 ppm (5). In the study presented here, tumors tooklonger to reach a maximum size when compared with the previousstudy, which was likely attributable to the lower concentrations ofgenistein in the diet. Cellular proliferation was increased in a dose-dependent manner as genistein concentrations increased in both thesoy protein and casein-based diets consistent with the tumor growthdata. The increase in MCF-7 cell proliferation was likely attributableto an estrogenic effect as indicated by the modest increase in pS2
Fig. 2. Average tumor area for week 29. Female ovariectomized athymic mice were
expression. These findings suggest that genistein, when fed to athymic
implanted with a 2-mg E pellet. The animals were then injected with 1.5 ϫ 105 MCF-7
mice in pure form or in a food source rich in the isoflavone such as
cells/site in four locations. Subsequently, tumors developed and were allowed to grow toan average cross-sectional area of 40 mm2. At this time, E pellets were removed from all
soy protein isolate, can stimulate estrogen-dependent tumor growth in
of the mice and they were assigned to one of eight treatment groups: positive controls that
were reimplanted with a new 2-mg E pellet, negative controls that were fed AIN 93G
rodent diet alone, LSI containing 15 ppm genistein, MSI containing 150 ppm genistein,
The estrogenic activity of isoflavones has been well documented in
HSI containing 300 ppm genistein, LG AIN 93G ϩ 15 ppm genistein, MG AIN
in vitro, in vivo, and clinical studies. Genistein can bind to the
93G ϩ 150 ppm genistein, and HG AIN 93G ϩ 300 ppm genistein. Tumors from each
estrogen receptor ␣ with an affinity 100-1000 times lower than E2
treatment group were evaluated by measuring tumor area. Measurements from all of thetumors in each treatment group were combined and averaged. Bars, SE.
(21). Additionally, in ovariectomized Sprague Dawley rats, dietary
SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
growth of MCF-7 cells in vitro (2, 5) and in vivo at 1 M (5). InMCF-7 cells, at concentrations from 20 –90 M genistein inhibitedDNA synthesis, but stimulated DNA synthesis at concentrations of0.1–10 M (31). In this study we demonstrated that dietary genisteinconsumed at 15 ppm, whether from soy protein or as a pure com-pound, does not stimulate growth that is significantly different fromthe negative control group lacking any form of estrogen exposure inregard to stimulation of MCF-7 tumor growth. These results suggestthat there is a level of dietary genistein that will not lead to an increasein tumor growth. Additional investigation will be necessary to deter-mine the concentration of this threshold dose.
Numerous studies have focused on the chemopreventative effects
of genistein and demonstrate that if exposed to isoflavones beforepuberty a rat has a reduced risk of developing breast cancer when
Fig. 4. Effect of soy and genistein on the cellular proliferation within MCF-7 tumors.
Tumors were removed from the mice for immunohistochemical analysis. Incorporation of
exposed to a chemical carcinogen (8 –10). It is reasonable then to
BrdUrd into cellular DNA was used as a marker of cellular proliferation. Immunohisto-
assume a similar protective effect might be seen in women if they are
chemistry was used to stain for cells containing BrdUrd. Positively staining as well as
exposed to isoflavones before puberty. Early exposure of women in
background cells were counted to give a final count on both proliferating and total cellsin a given area of tissue. Cell counts from each treatment group were then combined and
Asia to isoflavones may explain in part why Asian women have a
averaged (n ϭ 25 fields/five tumors/treatment group). The data are presented as the
lower rate of breast cancer incidence when compared with American
percentage of cells actively proliferating in a given area of tissue. Bars, SE.
women. It is hypothesized that a high soy diet throughout life protectsagainst development of breast cancer. This idea is supported by the
genistein (750 ppm) enhanced lobular-alveolar mammary gland de-
fact that upon moving to the United States, Asian women have a
velopment, increased uterine weight, and at 750 ppm, increased pitu-
similar breast cancer rate as that of American women after two
itary prolactin secretion and serum prolactin levels. In the uterus,
generations, indicating early exposure is critical (32–34). In evaluat-
dietary genistein increased c-fos mRNA expression (21). In vitro
ing the correlation between urinary phytoestrogen levels and breast
studies showing an increase in the levels of the estrogen-responsive
cancer incidence in postmenopausal women, a recent report showed
genes pS2 and c-fos when cells are treated with genistein, provide
high urinary genistein excretion was weakly and nonsignificantly
additional evidence of the estrogenic property of genistein (5, 22).
associated with a reduced breast cancer risk (35). Another study
Clinical studies have also confirmed the preclinical studies evaluating
shows no effect of soy consumption on the progression of chemically
the estrogenicity of genistein. Premenopausal women consuming tex-
induced mammary tumors in the rat (11). However, it is important to
tured vegetable protein containing 45 mg of isoflavones had pro-
note that in this study animals were intact with normal circulating E .
longed menstrual cycles. Additionally, the follicular phase was ex-
Paradoxically, we have shown that, in ovariectomized mice trans-
tended by suppression of the normal surge of follicle-stimulating
planted with human estrogen-dependent breast cancer cells, genistein
hormone and luteinizing hormone (23, 24). These data in conjunction
and now soy protein isolates containing genistein stimulated tumor
with the observations that dietary soy increases cell proliferation in
growth (5). Ovariectomized athymic mice implanted with MCF-7
human breast tissue (6) and increases pS2 expression (7) demonstrates
cells are an appropriate model of postmenopausal women with estro-
biological (estrogenic) activity in humans.
gen-dependent breast cancer. Plasma levels of E in ovariectomized
Isoflavones are being marketed to postmenopausal women in both
athymic mice are 27–38 pg/ml compared with postmenopausal
supplement form and in soy food products for the relief of the
women 10 – 40 pg/ml (36). We believe the low endogenous estrogen
symptoms of menopause. It is, therefore, critical to understand what
environment created in this model by ovariectomizing the mice
activities genistein may have in these women. Consideration must be
affords genistein the opportunity to be a significant source of estro-
given to how genistein may influence the endogenous levels of serum
genicity in these animals and, hence, allows for the compound to
E in women consuming the compound. There is evidence suggesting
stimulate the growth of these tumors. Therefore, if a postmenopausal
diets high in soy have the ability to lower serum E concentrations
woman with low endogenous E levels has an existing estrogen-
(25, 26). A study supporting this finding showed that normally cycling
dependent breast tumor it is also possible that consumption of dietary
women consuming a soy diet containing ϳ154 mg total isoflavones/
genistein from various food sources including soy protein isolate may
day had a 25% reduction in circulating E (27). Contrarily, other
produce sufficient plasma levels of genistein to result in enhanced
studies in which premenopausal women have consumed soy have
estrogen-dependent tumor growth as observed in athymic mice. These
found no change in serum E concentrations (6, 28), and one study
results collectively suggest that the time of exposure to genistein is
showed increased serum E in women consuming soy (23). It is
critical to the overall effect genistein will have on estrogen-dependent
unclear how isoflavones in soy effect normal circulating E concen-
trations in premenopausal women. Postmenopausal women consum-
From the discussion presented here it is clear that the isoflavones
ing a soy diet have been shown to have no change in serum circulating
possess diverse biological activities and potency. These activities are
E levels (28). This finding in conjunction with reports that genistein
often dependent upon the concentration and timing of administration
acts estrogenically, and postmenopausal women naturally have low
of the isoflavones. As a result, it is important to focus on what
plasma concentrations of E , suggest that in a postmenopausal woman,
activities occur at plasma concentrations that are relevant to what is
these weak estrogens may have significant estrogenic activity.
observed in humans consuming isoflavone containing diets. In regard
At concentrations above 10 M genistein has been demonstrated to
to breast cancer, the isoflavones, specifically genistein, have paradox-
inhibit in vitro cell proliferation in a variety of cell types including
ical effects that can be resolved when you consider dosage and timing
estrogen-dependent (MCF-7) and estrogen-independent (MDA-468)
of administration. For example, prepubertal exposure to genistein
human breast cancer cells (29). Genistein has also been shown to
appears to be protective against the development of breast cancer, but
inhibit activities of protein tyrosine kinase at concentrations Ͼ20 M
consumption of the phytoestrogen in either pure form or in soy protein
(30). However, at levels as low as 200 nM, genistein stimulates the
isolate, after development of an estrogen-dependent breast cancer may
SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
enhance the growth of that tumor as determined by this study. Addi-
and provide an in vivo model to assess the responsiveness of cells to estrogen.
tional investigation into the biological activities of the isoflavones and
Biochem. Pharmacol., 43: 1263–1267, 1992.
16. Gottardis, M. M., Jiang, S. Y., Jeng, M. H., and Jordan, V. C. Inhibition of
soy itself is needed. The use of soy protein isolate as a dietary
tamoxifen-stimulated growth of an MCF-7 tumor variant in athymic mice by novel
supplement may have numerous health benefits such as reducing risk
steroidal antiestrogens. Cancer Res., 49: 4090 – 4093, 1989.
17. Helferich, W. G., Jump, D. B., Anderson, D. B., Skjaerlund, D. M., Merkel, R. A., and
of coronary heart disease by lowering serum cholesterol. However, for
Bergen, W. G. Skeletal muscle ␣-actin synthesis is increased pretransitionally in pigs
the subgroup of postmenopausal women who have or are at high risk
fed the phenethanolamine ractopamine. Endocrinology, 126: 3096 –3100, 1990.
of developing breast cancer there is need for additional consideration
18. Brown, A. M. C., Jeltsch, J. M., Roberts, M., and Chambon, P. Activation of pS2 gene
transcription is a primary response to estrogen in the human breast cancer cell line
into the possibility of enhancing estrogen-dependent tumor growth by
MCF-7. Proc. Natl. Acad. Sci. USA, 81: 6344 – 6348, 1984.
consumption of isoflavone containing products.
19. De Fazio, A., Leary, J. A., Hedley, D. W., and Tattersall, M. H. Immunohistochemical
detection of proliferating cells in vivo. J. Histochem. Cytochem., 35: 571–577, 1987.
20. Spady, T. J., Harwell, D. M. E., Snyder, M. C., Pennington, K. L., Mcomb, R. D., and
REFERENCES
Shull, J. D. Estrogen-induced tumorigenesis in the Copenhagen rat: disparate suscep-tibilities to development of prolactin producing pituitary tumors and mammary
1. Soy protein and coronary heart disease. Federal Register., 64: 57699 –57733, 1999.
carcinomas. Cancer Lett., 124: 95–103, 1998.
2. Martin, P. M., Horwitz, K. B., Ruyan, D. S., and McGuire, W. L. Phytoestrogen
21. Santell, R. C., Chang, Y. C., Nair, M. G., and Helferich, W. G. Dietary genistein
interaction with estrogen receptors in human breast cancer cells. Endocrinology, 103:
exerts estrogenic effects on the uterus, mammary gland, and the hypothalamic/
pituitary axis in rats. J. Nutr., 127: 263–269, 1997.
3. Setchell, K., Borriello, S. P., Hulme, P., Kirk, D. N., and Axelson, M. Nonsteroidal
22. Sathyamoorthy, N., Wang, T. T. Y., and Phang, J. M. Stimulation of pS2 expression
estrogens of dietary origin: possible roles in hormone-dependent disease. Am. J. Clin.
by diet-derived compounds. Cancer Res., 54: 957–961, 1994.
23. Cassidy, A., Bingham, S., and Setchell, K. Biological effects of a diet of soy protein
4. Farmakalikis, E., Hathcock, J. N., and Murphy, P. A. Oestrogenic potency of genistin
rich in isoflavones on the menstrual cycle of premenopausal women. Am. J. Clin.
and daidzin in mice. Food Chem. Toxic., 23: 741–745, 1985.
5. Hsieh, C. Y., Santell, R. C., Haslam, S. Z., and Helferich, W. G. Estrogenic effects
24. Cassidy, A., Bingham, S., and Setchell, K. Biological effects of isoflavones in young
of genistein on the growth of estrogen receptor-positive human breast cancer (MCF-7)
women: importance of the chemical composition of soyabean products. Br. J. Nutr.,
cells in vitro and in vivo. Cancer Res., 58: 3833–3838, 1998.
6. Petrakis, N. L., Barnes, S., King, E. B., Lowenstein, J., Wiencke, J., Lee, M. N.,
25. Nagata, C., Takatsuka, N., Inaba, S., Kawakami, N., and Shimizu, H. Effect of soy
Miike, R., Kirk, M., and Coward, L. Stimulatory influence of soy protein isolate on
milk consumption on serum estrogen concentrations in premenopausal Japanese
breast fluid secretion in pre- and postmenopausal women. Cancer Epidemiol. Bi-
women. J. Natl. Cancer Inst., 90: 1830 –1835, 1998.
omark. Prev., 5: 785–794, 1996.
26. Xu, X., Duncan, A. M., Merz, B. E., and Kurzer, M. S. Effects of soy isoflavones on
7. Hargreaves, D. F., Potten, C. S., Harding, C., Shaw, L. E., Morton, M. S., Roberts,
estrogen and phytoestrogen metabolism in premenopausal women. Cancer Epidemiol.
S. A., Howell, A., and Bundred, N. J. Two-week dietary soy supplementation has an
Biomark. Prev., 12: 1101–1108, 1998.
estrogenic effect on normal premenopausal breast. J. Clin. Endocrinol. Metab., 84:
27. Lu, L. W., Anderson, K. E., Grady, J. J., Kohen, F., and Nagamani, M. Decreased
ovarian hormones during a soya diet: implications for breast cancer prevention.
8. Fritz, W. A., Coward, L., Wang, J., and Lamartiniere, C. A. Dietary genistein:
Cancer Res., 60: 4112– 4121, 2000.
perinatal mammary cancer prevention, bioavailability and toxicity testing in the rat.
28. Baird, D. D., Umbach, D. M., Lansdell, L., Hughes, C. L., Setchell, K. D. R.,
Carcinogenesis (Lond.), 19: 2151–2158, 1998.
Weinberg, C. R., Haney, A. F., Wilcox, A. J., and McLachlan, J. A. Dietary
9. Lamartiniere, C. A., Moore, J. B., Brown, N. M., Thompson, R., Hardin, J. J., and
intervention study to assess estrogenicity of dietary soy among postmenopausal
Barnes, S. Genistein suppresses mammary cancer in rats. Carcinogenesis (Lond.), 16:
women. J. Clin. Endocrinol. Metab., 80: 1685–1690, 1995.
29. Peterson, G., and Barnes, S. Genistein inhibition of the growth of human breast cancer
10. Murrill, W. B., Brown, N. M., Zhang, J. X., Manzolillo, P. A., Barnes, S., and
cells: independence from the estrogen receptor and the multi-drug resistance gene.
Lamartiniere, C. A. Prepubertal genistein exposure suppresses mammary cancer and
Biochem. Biophys. Res. Comm., 179: 661– 667, 1991.
enhances gland differentiation in rats. Carcinogenesis (Lond.), 17: 1451–1457, 1996.
30. Peterson, G. Evaluation of the biochemical targets of genistein in tumor cells. J. Nutr.,
11. Cohen, L. A., Zhao, Z., Pittman, B., and Scimeca, J. A. Effect of intact and
isoflavone-depleted soy protein on NMU-induced rat mammary tumorigenesis. Car-
31. Wang, C., and Kurzer, M. Phytoestrogen concentration determines effects on DNA
cinogenesis (Lond.), 21: 929 –935, 2000.
synthesis in human breast cancer cells. Nutr. Cancer, 28: 236 –247, 1997.
12. Xu, X., Wang, H. J., Murphy, P. A., Cook, L., and Hendrich, S. Daidzein is a more
32. Buell, P. Changing incidence of breast cancer in Japanese-American women. J. Natl.
bioavailable soymilk isoflavone than is genistein in adult women. J. Nutr., 124:
Cancer Inst., 51: 1479 –1483, 1973.
33. Hirayama, T. Epidemiology of breast cancer with special reference to the role of the
13. Reeves, P. G., Nielsen, F. H., and Fahey, G. C., Jr. AIN-93 purified diets for
diet. Prev. Med., 7: 173–195, 1978.
laboratory rodents: final report of the American Institute of Nutrition ad hoc writing
34. Kodama, M., Kodama, T., Miura, S., and Yoshida, M. Nutrition and breast cancer risk
committee on the reformulation of AIN -76A rodent diet. J. Nutr., 123: 1939 –1951,
in Japan. Anticancer Res., 11: 745–754, 1991.
35. Tonkelaar, I., Keinan-Boker, L., Van’t Veer, P., Arts, C. J. M., Adlercreutz, H.,
14. McManus, M. J., and Welsch, C. W. Hormone-induced ductal DNA synthesis of
Thijssen, H. H., and Peeters, H. M. Urinary phytoestrogens and postmenopausal
human breast tissues maintained in the athymic nude mouse. Cancer Res., 41:
breast cancer risk. Cancer Epidemiol. Biomarkers Prev., 10: 223–228, 2001.
36. Seibert, K., Shafie, S. M., Triche, T. J., Whang-Peng, J. J., O’Brien, S. J., Toney,
15. Noel, A., Simon, N., Raus, J., and Foidart, J. M. Basement membrane components
J. H., Huff, K. K., and Lippman, M. E. Clonal variation of MCF-7 breast cancer cells
(Matrigel) promote the tumorigenicity of human breast adenocarcinoma MCF7 cells
in vitro and in athymic nude mice. Cancer Res., 43: 2223–2239, 1983.
Writ Petition No. 4361 of 2009 In the matter of: An application under Article 102 of the Constitution of the People’s Republic of Bangladesh. In the matter of: Sheikh Md. Romij Uddin (Tara Miah) and another Mr. Majedul Islam Patwary with Mr. Syed Hasan Zahir, Advocates Heard on 28.9.2010 and Judgment on 04.10.2010 Present: Ms. Justice Nazmun Ara Sultana And Mr. Justi
Haloperidol: MedlinePlus Drug Information Haloperidol URL of this page: http://www.nlm.nih.gov/medlineplus/druginfo/meds/a682180.html IMPORTANT WARNING: Studies have shown that older adults with dementia (a brain disorder that affects the ability to remember, think clearly,communicate, and perform daily activities and that may cause changes in mood and personality) who takeantipsychotics (

 SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
with RNase-free dH O. RNA was stored at Ϫ80°C. RNA concentration was
ϭ 40 g of single-stranded RNA/ml).
SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
with RNase-free dH O. RNA was stored at Ϫ80°C. RNA concentration was
ϭ 40 g of single-stranded RNA/ml).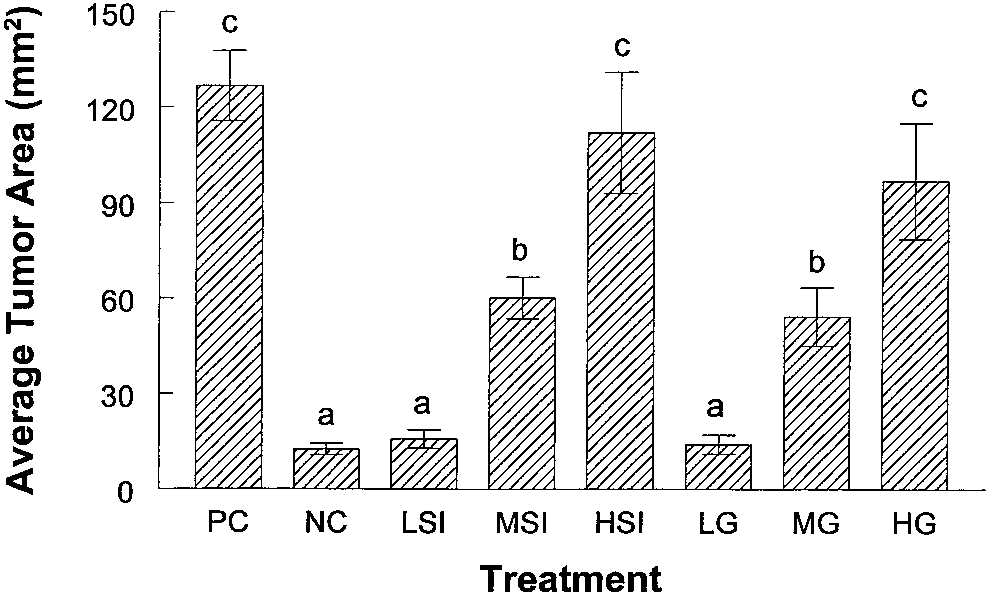
 SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
(Fig. 2). Body weight was monitored weekly, and no significantdifference was observed among the treated and control groups (datanot shown). These data indicate that dietary soy isolates containingincreasing concentrations of genistein and casein-based diets contain-ing equalized concentrations of genistein act in a dose-dependentmanner to stimulate growth of human estrogen-dependent breastcancer cells transplanted into athymic mice. Also, it is important tonote that there were no significant differences in growth of tumors inthe negative control, low soy isolate, and low genistein groups. Thesedata suggest that there is a threshold level of dietary genistein belowwhich no increase in estrogen-dependent tumor growth is observed.
SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
(Fig. 2). Body weight was monitored weekly, and no significantdifference was observed among the treated and control groups (datanot shown). These data indicate that dietary soy isolates containingincreasing concentrations of genistein and casein-based diets contain-ing equalized concentrations of genistein act in a dose-dependentmanner to stimulate growth of human estrogen-dependent breastcancer cells transplanted into athymic mice. Also, it is important tonote that there were no significant differences in growth of tumors inthe negative control, low soy isolate, and low genistein groups. Thesedata suggest that there is a threshold level of dietary genistein belowwhich no increase in estrogen-dependent tumor growth is observed.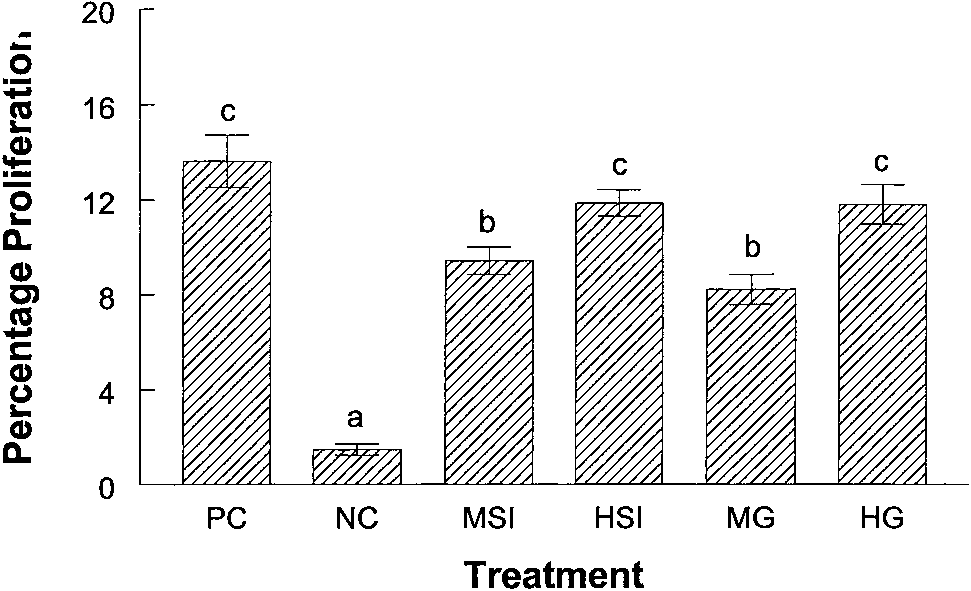 SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
growth of MCF-7 cells in vitro (2, 5) and in vivo at 1 M (5). InMCF-7 cells, at concentrations from 20 –90 M genistein inhibitedDNA synthesis, but stimulated DNA synthesis at concentrations of0.1–10 M (31). In this study we demonstrated that dietary genisteinconsumed at 15 ppm, whether from soy protein or as a pure com-pound, does not stimulate growth that is significantly different fromthe negative control group lacking any form of estrogen exposure inregard to stimulation of MCF-7 tumor growth. These results suggestthat there is a level of dietary genistein that will not lead to an increasein tumor growth. Additional investigation will be necessary to deter-mine the concentration of this threshold dose.
SOY PROTEIN AND E2-DEPENDENT BREAST CANCER TUMOR GROWTH
growth of MCF-7 cells in vitro (2, 5) and in vivo at 1 M (5). InMCF-7 cells, at concentrations from 20 –90 M genistein inhibitedDNA synthesis, but stimulated DNA synthesis at concentrations of0.1–10 M (31). In this study we demonstrated that dietary genisteinconsumed at 15 ppm, whether from soy protein or as a pure com-pound, does not stimulate growth that is significantly different fromthe negative control group lacking any form of estrogen exposure inregard to stimulation of MCF-7 tumor growth. These results suggestthat there is a level of dietary genistein that will not lead to an increasein tumor growth. Additional investigation will be necessary to deter-mine the concentration of this threshold dose.