Sex Differences in Salivary Cortisol Levels Following Naltrexone Administration1 Department of Biobehavioral Health
LARRY D. JAMNER, JANEL ALBERTS, MATTHEW D. ORENSTEIN,
University of California, San FranciscoFresno Veterans Affairs Medical Center
Effects of endogenous opioid peptide blockade by naltrexone on salivary cortisol levelswere examined in healthy men (n = 8) and women (n = 6). Participants received naltrex-one (100 mg) during one laboratory session and a placebo pill during another session. Drug order was counterbalanced across participants. Saliva samples were collected 24 hrafter each pill was administered. Among women, salivary cortisol levels significantlyincreased following naltrexone administration compared with a placebo pill. Naltrexoneadministration did not alter salivary cortisol levels in men. Results suggest sex differencesin neuroendocrine sensitivity to opioid blockade, a finding that may hold significance withregard to the treatment of alcohol addiction with naltrexone.
The endogenous opioid peptide system has several physiological roles includ-
ing pain inhibition, appetite regulation, and modulation of the stress response
1This work was conducted at the University of California, Irvine, and was supported partially by
a University of California, Irvine, career development award (447614-09503-8) granted to the firstauthor. This study was supported partially by a Merit Review grant awarded to H. Leigh by theDepartment of Veterans Affairs. Additional support for biochemical assays was provided by NIMHgrant 5T32MH19958-03. The procedures followed in this research were reviewed and approved byUniversity of California, Irvine, human-subjects committees and were performed in accordance withhuman-use ethical standards. These findings were presented at the 38th meeting of the Society forPsychophysiological Research. We thank Larry Cahill, Elissa Epel, and Douglas Granger for theirassistance, as well as William Lovallo and Regan A. R. Gurung for their comments.
2Correspondence concerning this article should be addressed to Laura Cousino Klein, Depart-
ment of Biobehavioral Health, Penn State University, 315 East Health and Human Development, Uni-versity Park, PA 16802-6509. e-mail: LXK18@psu.edu
Journal of Applied Biobehavioral Research, 2000, 5, 2, pp. 144-153. Copyright 2000 by Bellwether Publishing, Ltd. All rights reserved.
through the hypothalamic-pituitary-adrenomedullary (HPA) axis. Endogenousopioid peptides (EOPs) are derived from proopiomelanocortin (POMC), whichalso is the precursor for adrenocorticotropin hormone (ACTH). Both ACTH andEOPs are released from the anterior pituitary, and ACTH stimulates release ofglucocorticoids (e.g., cortisol) from the adrenal glands. EOP blockade by opioidantagonists (e.g., naloxone and naltrexone) also has important effects on HPA-axis functioning.
Earlier studies report that the administration of naloxone, a short-acting opi-
oid antagonist used in the treatment of opioid overdose, increases plasma cortisollevels in men (Grevert, Albert, Inturrisi, & Goldstein, 1983; Grossman, Gaillard,McCartney, Rees, & Besser, 1982; Grossman, Moult, Cunnah, & Besser, 1986). It is believed that naloxone increases cortisol release by suppressing theACTH inhibitory effects of EOPs in humans (Grossman et al., 1982; Morley,Baranetsky, Wingert, Carlson, & Hershman, 1980; Volvavka, Cho, Mallya, &Bauman, 1979). Our review of the literature on humans suggests that the reportedeffects of opioid antagonism on HPA-axis activity are based primarily on theadministration of naloxone, which produces short-term opioid receptor blockade(i.e., 90 min). It is possible that the long-term blockade of endogenous opioidreceptors has different physiological consequences than does the acute, short-term blockade by naloxone.
Naltrexone is a long-acting opioid antagonist that works primarily on µ-
opioid receptors to block the binding of EOPs (e.g., β-endorphin) and exogenousopiates (e.g., morphine) to these receptors (Jaffe & Martin, 1990). Naltrexone hasa plasma half-life of about 10 hr (depending on dose administration), and it isextensively metabolized into the primary active metabolite, 6-β-naltrexol, by theliver. Because of the pharmacokinetic properties of naltrexone and its metabo-lites, a single 100-mg oral dose of naltrexone will block µ-opioid receptors for atleast 48 hr (Way, Fields, & Way, 1998). One study indicated that µ-opioid recep-tors in the brain remained blocked up to 100 hr after the administration of a doseas low as 50 mg/day (Lee et al., 1988). This point is noteworthy given that naltr-exone recently received Food and Drug Administration (FDA) approval as apharmacologic treatment for alcoholism. In these clinical cases, men and womenare dosed daily with 50 to 100 mg of naltrexone, over a course of several months,to curb alcohol cravings and decrease alcohol consumption (for review, seeWeinrieb & O’Brien, 1997). Although this antagonist is used as a long-term treat-ment for alcoholism (Litten & Allen, 1998, 1999), little is known about the long-term effects of endogenous opioid blockade on HPA-axis activity in healthy vol-unteers. Empirical studies on the HPA-axis effects of naltrexone examine neu-roendocrine changes for up to 12 hr after drug administration; clinically,however, naltrexone is administered in doses designed to block µ-opioid recep-tors for up to 48 to 72 hr (depending on dose) to treat alcoholism. Our review ofthe literature found only one published report of the effects of naltrexone on
Demographic Characteristics of Sample (Means ± Standard Error of Mean)
neuroendocrine function in humans (Mendelson, Ellingboe, Keuhnle, & Mello,1979). However, naltrexone’s effect on cortisol levels was not examined, andwomen were not included in the study. Therefore, the present study examined theeffects of naltrexone administration on salivary cortisol (i.e., free cortisol)responses 24 hr later in men and women as a first step to understanding thelonger time course of the HPA-axis effects of naltrexone administration.
Participants included 16 healthy nonsmokers (8 women and 8 men) between
the ages of 18 and 45 (mean = 31.56 years) who were recruited from the Univer-sity of California, Irvine, campus and local community through e-mail and flyersas part of a larger ongoing project investigating the effects of naltrexone onambulatory physiological functioning. The men and women were similar in age,years of education, body mass index, ethnicity, and marital status (Table 1). Uponscreening, individuals who reported health problems, histories of alcohol or drugabuse, or current medication use were excluded. Volunteers were paid for theirparticipation in the larger project, which lasted for 3 days.
Menstrual cycle phase was not controlled for in the present experiment, and
women on oral contraceptives (OC) also were included in the study. Although
menstrual cycle and OC use influence salivary free cortisol responses to stress inwomen, basal salivary free cortisol levels (i.e., free cortisol levels upon awaken-ing in the morning) do not differ among women in the luteal or follicular phasesof the menstrual cycle or among those using oral contraceptives (Kirschbaum,Kudielka, Gaab, Schommer, & Hellhammer, 1999).
The study incorporated a within-subject double-blind randomized design. In
one session, participants ingested an oral dose (100 mg) of naltrexone hydro-chloride (Trexan®; DuPont Pharma, Delaware). In the other session, theyingested an inert placebo pill (DuPont Pharma, Delaware). The naltrexone dosewas selected on the basis of current research that indicates effective opioid block-ade in male and female alcohol-dependent patients at doses ranging from 20 mg/day to 100 mg/day (Litten & Allen, 1998; Way et al., 1998). When naltrexonedoses were adjusted for body weight (kg), women received 1.61 ± 0.04 mg nal-trexone/kg and men received 1.29 ± 0.07 mg naltrexone/kg. Drug conditionswere counterbalanced across participants.
The participants completed an orientation session that was followed by two
drug sessions, which were 1 week apart. During the orientation and afterinformed consent was received, the participants were administered a health-history interview to ensure their eligibility for the study, which included the stip-ulation that they have no history of alcohol or substance abuse. In addition, ablood sample was drawn and assayed for healthy liver enzyme activity. Womenwere given pregnancy tests at the beginning of each experimental session toensure that they were not pregnant before receiving the medication.
The participants arrived at the lab where they were given a pill to swallow
with a glass of water. They were observed in the lab for 90 min, and then theywere asked to come back the following morning to complete a variety of tasks aspart of a larger study. The participants were asked to abstain from prescription orover-the-counter medications, alcohol, and nicotine products 24 hr before arriv-ing at the lab and for 3 days after consuming the pill. The participants also wereasked to limit their caffeine intake to two servings of caffeine per day (i.e., two8 oz. cups of coffee or caffeinated sodas).
Saliva samples were collected in the laboratory the next morning when
participants arrived for the laboratory session. Samples were collected 24 hr
after each pill was administered (i.e., naltrexone and placebo), between 8:00 a.m. and 10:30 a.m. For each participant, collection times were similar across sessionsto control for potential circadian changes in cortisol levels (Grossman et al.,1982; Ockenfels et al., 1995). Salivary cortisol levels were assessed 24 hr afterthe pills were administered to examine the long-term effects of endogenous opi-oid blockade. Previous studies indicate that a 100-mg dose of naltrexone resultsin µ-opioid receptor blockade for up to 100 hr after administration (Lee et al.,1988).
After relaxing and completing questionnaires about the previous 24-hr
period, the participants were asked to produce a saliva sample by chewing a pieceof gum (Trident™ sugarless spearmint gum) for 2 min (Schwartz, Granger,Susman, Gunnar, & Laird, 1998). Next, they were asked to roll a cotton swab intheir mouths for an additional 2 min before being instructed to place the swabinto a Salivette® saliva collection tube (Sarstedt, Inc., North Carolina). Samplesimmediately were placed on ice and then frozen (-70 degrees Celsius) for laterassessment of cortisol by radioimmunoassay (Coat-a-Count® assay; DiagnosticProducts Corporation, Los Angeles, California). Samples were shipped on dryice by overnight delivery to the Behavioral Endocrinology Laboratory at ThePennsylvania State University, University Park campus, where the cortisol assayswere conducted. All samples were tested in duplicate in a minimum number ofassay batches. Values used in data analyses were the averages of the duplicatetests. The assay had a lower limit of sensitivity of 0.03 µg/dl, with an averageinter- and intra-assay covariance of less than 10% and 5%, respectively. Individ-ual samples with duplicate tests that varied by more than 5% were repeated insubsequent runs. Treatment of Data and Statistical Analyses
Two samples (1 placebo day and 1 naltrexone day) from two different female
participants were lost during sample processing. Therefore, these women are notincluded in the analyses. Natural logarithmic transformations were applied tothe raw data because the raw cortisol values were not normally distributed. Because of the variability in cortisol responses, previous investigations of sali-vary cortisol have applied logarithmic transformations to salivary cortisol data(e.g., Ockenfels et al., 1995). In the present study, this transformation resulted ina normal distribution of the data; therefore, all reported analyses are based onthese logarithmic-transformed values. However, raw cortisol values (± standarderror of the mean) were used to graph the data for clarity.
Because women received a greater naltrexone dosage than did men, F(1,
13) = 11.87, p < .05, the following analyses were conducted using dosage as acovariate. However, naltrexone dosage did not emerge as a significant covariatein any of the analyses; therefore, analyses without the covariate are reported. Figure 1. Effects of naltrexone on salivary cortisol levels (µg/dl) in men and women.
Error bars are equal to the standard error of the mean.
Figure 1 presents mean salivary cortisol levels following naltrexone and
placebo administration in men and women. A repeated-measures analysis ofvariance (ANOVA) was conducted to evaluate salivary cortisol responses to nal-trexone compared with placebo cortisol levels. The results revealed a significantdrug by sex interaction, F(1, 12) = 4.71, p = .05. Specifically, among women, sal-ivary cortisol levels were significantly higher in response to naltrexone comparedwith the placebo condition, F(1, 5) = 7.04, p < .05. In contrast, naltrexone admin-istration did not alter salivary cortisol levels in men compared with the placebocondition. Separate ANOVAs indicated that there were no sex differences in cor-tisol levels under naltrexone or placebo conditions.
The present study examined the effects of endogenous opioid blockade by
naltrexone on salivary cortisol responses in men and women. The results suggestthat naltrexone administration significantly increases salivary cortisol 24 hr afteradministration in women but not in men. This finding for men is surprising inlight of previous reports of increases in plasma cortisol responses among menshortly following administration of naloxone and naltrexone (Grevert et al.,1983; Grossman et al., 1982, 1986).
There are several pharmacokinetic and pharmacodynamic explanations
for why the present results may differ from the current literature. Althoughpharmacologically similar to naloxone, naltrexone is two to eight times more
potent than naloxone (Blumberg & Dayton, 1973; Verebely & Mule, 1975), andnaltrexone’s actions last hours longer than do naloxone’s actions (Martin, Jasin-ski, & Mansky, 1973). It is possible that long-term blockade of endogenous opi-oid receptors resulted in an attenuation or habituation of the cortisol responseamong men but not among women, suggesting a different dose-response curvefor men and women. A recent study observed higher µ-opioid receptor bindingamong women (Zubieta, Dannals, & Frost, 1999), suggesting that the observedincrease in cortisol in the present study was a result of greater antagonism of thecentral opioid system stemming from increased naltrexone binding in women. These results add to an increasing body of literature on sex differences in opioidsensitivity and opioid-hormone interactions in humans and animals (e.g.,Barbarino et al., 1987; Crowley, 1988; Fernandez et al., 1999; Klein, Popke, &Grunberg, 1998; Kreek, Schluger, Borg, Gunduz, & Ho, 1999; Schlenker, Martin,Lin, & Egland, 1997; Zubieta et al., 1999). Interestingly, there are no publishedreports on sex differences in the pharmacokinetics of naltrexone per se, and thenaltrexone manufacturer, DuPont Pharma (Wilmington, DE), reports no indica-tion of gender differences in the pharmacokinetics of naltrexone in companyrecords (L. Lupo, personal communication, November 14, 2000). Thus, the issuesurrounding sex differences in naltrexone metabolism, distribution, absorption,and excretion, as well as the subsequent impact on neuroendocrine functioning,remains unclear. The present results suggest that future studies need to directlyaddress this question.
Another point directly related to understanding the observed sex difference in
cortisol responses to naltrexone is that perhaps higher naltrexone dosages amongwomen contributed to the observed increase in cortisol levels. We addressed thisquestion by conducting a separate analysis of covariance (ANCOVA) on cortisollevels with mean naltrexone dosage (i.e., mg naltrexone/kg body weight) as thecovariate, and we found that the main effect for naltrexone on cortisol levelsremained. Although naltrexone is administered clinically in 50- or 100-mg dosesto patients without adjusting for body weight, a direct comparison of cortisolresponses to naltrexone in men and women needs to be made with naltrexoneamounts adjusted for body weight.
A final explanation for the differences between previously reported results
and the present findings is that cortisol levels were assessed 24 hr after theadministration of naltrexone. No other studies have evaluated these long-termeffects of opioid blockade on HPA-axis activity. These findings indicate that, atthe dose administered, women demonstrate greater HPA-axis sensitivity toendogenous opioid blockade than do men. Given that this dose (100 mg) is simi-lar to that used in alcohol addiction treatment, this finding may have importantimplications for chronic alcohol treatment with naltrexone.
The present results should be approached with caution, however, because of
the small sample size and single saliva sample collection procedures. The use of
a single saliva sample method prevents an assessment of time effects on cortisolresponses beyond the influence of naltrexone. Furthermore, situational factorscould account for cortisol differences between men and women, but they werenot captured because of the small window of cortisol assessment. These limita-tions are important in light of the relevance of the present findings to understand-ing interactions between the HPA-axis and EOP systems. The present experimentis a first step in developing an understanding of the time-course neuroendocrineeffects of naltrexone administration. The results suggest that sex differences maycontribute to naltrexone’s efficacy in the treatment of alcoholism. They also indi-cate that additional investigations on sex differences in naltrexone’s neuroendo-crine effects are necessary.
Barbarino, A., De Marinis, L., Mancini, A., D’Amico, C., Passeri, M., Zuppi, P.,
Sambo, P., & Tofani, A. (1987). Sex-related naloxone influence on growth hormone-releasing hormone-induced growth hormone secretion in normal subjects. Metabolism, 36, 105-109.
Blumberg, H., & Dayton, H. B. (1973). Naloxone, naltrexone, and related noroxy-
morphones. Advances in Biochemistry and Psychopharmacology, 8, 33-43.
Crowley, W. R. (1988). Sex differences in the responses of hypothalamic lutein-
izing hormone-releasing hormone and catecholamines systems to ovarian hormones and naloxone: Implications for sexual differentiation of luteinizing hormone secretion in rats. Brain Research, 461, 314-321.
Fernández, B., Antelo, M. T., Guaza, C., Alberti, I., Pinillos, M. L., & Viveros,
M. P. (1999). Naltrindole administration during the preweanling period and manipulation affect adrenocortical reactivity in young rats. Developmental Brain Research, 112, 135-137.
Grevert, P., Albert, L. H., Inturrisi, C. E., & Goldstein, A. (1983). Effects of
eight-hour naloxone infusions on human subjects. Biological Psychiatry,18, 1375-1392.
Grossman, A., Gaillard, R. C., McCartney, P., Rees, L. H., & Besser, G. M.
(1982). Opiate modulation of the pituitary-adrenal axis: Effects of stress and circadian rhythm. Clinical Endocrinology, 17, 279-286.
Grossman, A., Moult, P. J. A., Cunnah, D., & Besser, G. M. (1986). Different
opioid mechanisms are involved in the modulation of ACTH and gonadotro- phin release in man. Neuroendocrinology, 42, 357-360.
Jaffe, J. H., & Martin, W. R. (1990). Opioid analgesics and antagonists. In A. G.
Gilman, T. W. Rall, A. S. Nies, P. Taylor (Eds.), Goodman and Gilman's: Thepharmacological basis of therapeutics (8th ed., pp. 485-521). New York, NY:McGraw Hill.
Kirschbaum, C., Kudielka, B. M., Gaab, J., Schommer, N. C., & Hellhammer,
D. H. (1999). Impact of gender, menstrual cycle phase, and oral contracep- tives on the activity of the hypothalamus-pituitary-adrenal axis. Psycho- somatic Medicine, 61, 154-162.
Klein, L. C., Popke, E. J., & Grunberg, N. E. (1998). Sex differences in effects of
opioid blockade on stress-induced freezing behavior. Pharmacology, Bio- chemistry, & Behavior, 61, 413-417.
Kreek, M. J., Schluger, J., Borg, L., Gunduz, M., & Ho, A. (1999). Dynorphin
A1-13 causes elevation of serum levels of prolactin through an opioid receptor mechanism in humans: Gender differences and implications for modulation of dopaminergic tone in the treatment of addictions. The Journal of Pharma- cology and Experimental Therapeutics, 288, 260-269.
Lee, M. C., Wagner, H. N., Tanada, S., Frost, J. J., Bice, A. N., & Dannals, R. F.
(1988). Duration of occupancy of opiate receptors by naltrexone. Journal of Nuclear Medicine, 29, 1207-1211.
Litten, R. Z., & Allen, J. P. (1999). Medications for alcohol, illicit drug, and
tobacco dependence. An update of research findings. Journal of Substance Abuse Treatment, 16, 105-112.
Litten, R. Z., & Allen, J. P. (1998). Advances in development of medications for
alcoholism treatment. Psychopharmacology, 139, 20-33.
Martin, W. R., Jasinski, D. R., & Mansky, P. A. (1973). Naltrexone: An antago-
nist for the treatment of heroin dependence. Archives of General Psychiatry, 28, 784-791.
Mendelson, J. H., Ellingboe, J., Keuhnle, J. C., & Mello, N. K. (1979). Effects of
naltrexone on mood and neuroendocrine function in normal adult males. Psy- choneuroendocrinology, 3, 231-236.
Morley, J. E., Baranetsky, N. G., Wingert, T. D., Carlson, H. E., & Hershman,
J. M. (1980). Endocrine effects of naloxone-induced opiate receptor block- ade. Journal of Clinical Endocrinology and Metabolism, 50, 251-257.
Ockenfels, M. C., Porter, L., Smyth, J., Kirschbaum, C., Hellhammer, D. H., &
Stone, A. A. (1995). Effect of chronic stress associated with unemployment in salivary cortisol: Overall cortisol levels, diurnal rhythm, and acute stress reactivity. Psychosomatic Medicine, 57, 460-467.
Schlenker, E. H., Martin, D. S., Lin, X. M., & Egland, M. C. (1997). Naloxone
microinjected into the arcuate nucleus has differential effects on ventilation in male and female rats. Physiology & Behavior, 62, 531-536.
Schwartz, E. B., Granger, D. A., Susman, E. J., Gunnar, M. R., & Laird, B.
(1998). Assessing salivary cortisol in studies of child development. Child Development, 69, 1503-1513.
Verebely, K., & Mule, S. J. (1975). Naltrexone pharmacology, pharmacokinetics,
and metabolism: Current status. American Journal of Drug and Alcohol Abuse, 2, 357-363.
Volvavka, J., Cho, D., Mallya, A., & Bauman, L. (1979). Naloxone increases
ACTH and cortisol levels in man. New England Journal of Medicine, 300, 1056-1057.
Way, W. L., Fields, H. L., & Way, E. L. (1998). Opioid analgesics and antago-
nists. In B. G. Katzung (Ed.), Basic and clinical pharmacology (7th ed.,pp. 496-515). New York, NY: Appleton & Lange.
Weinrieb, R. M., & O’Brien, C. P. (1997). Naltrexone in the treatment of alcohol-
ism. Annual Review of Medicine, 48, 477-487.
Zubieta, J., Dannals, R. F., & Frost, J. J. (1999). Gender and age influences on
human brain mu-opioid receptor binding measured by PET. American Jour- nal of Psychiatry, 156, 842-848.
Swine Flu and the Great Flu Pandemic of 1918-19 The Similarities and What History Can Teach Us By Molly Punzo, M.D. The Great Flu Pandemic of 1918-19 killed more people than any other outbreak of disease in history. It is estimated that between 50 and 100 million died from what was then known as “Spanish Flu”. It most often killed those in the prime of life, and it killed with extra
Science citation index (SCI) publications:1. Triangular CdS nanocrystals: Structural and optical studiesN. Pinna, K. Weiss, J. Urban, M. P. PileniAdv. Mater. 2001, 13, 2612. Triangular CdS nanocrystals: Synthesis, characterization, and stabilityN. Pinna, K. Weiss, H. Sack-Kongehl, W. Vogel, J. Urban, M.P. PileniLangmuir, 2001, 17, 79823. Optical properties of silver nanocrystals self-organize
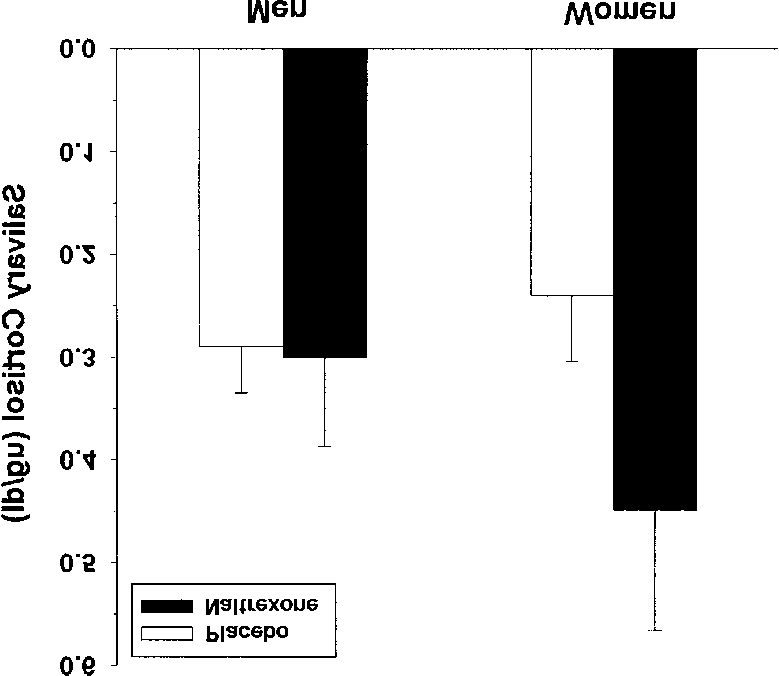 Figure 1. Effects of naltrexone on salivary cortisol levels (µg/dl) in men and women.
Figure 1. Effects of naltrexone on salivary cortisol levels (µg/dl) in men and women.