Demographic and genetic structures of white sea bream
populations (Diplodus sargus, Linnaeus, 1758) inside and outside
Laboratoire d’ichtyoécologie tropicale et méditerranéenne, École pratique des hautes études, CNRS UMR 8046,université de Perpignan, 52, av. de Villeneuve, 66860 Perpignan cedex, France
Received 6 September 2002; accepted 11 June 2003
Abstract
We studied the white sea bream (Diplodus sargus), a protandrous hermaphroditic fish, in two protected and unprotected areas
in southwestern France. We observed a significant difference in the demographic structure between the two areas. Females were present in two different age distributions inside and outside the marine reserve with younger females outside. This suggests plasticity in the age of sexual inversion in the case of an exploited population. Genetic differentiation was weak and apparent at only one locus of 26 surveyed (FST = 0.007, p = 0.04). Our data suggest that gene flow between the two areas is important, or the separation between the two sites is recent. Our data on the white sea bream show that fishes inside and outside the marine reserve are very similar genetically, which means that the ‘reserve effect’ is truly a demographic one, not the result of genetic differences. To cite this article: P. Lenfant, C. R. Biologies 326 (2003). 2003 Académie des sciences. Published by Éditions scientifiques et médicales Elsevier SAS. All rights reserved. Résumé Structures démographique et génétique des populations de sar commun (Diplodus sargus, Linnaeus, 1758) au sein et à l’extérieur d’une réserve marine en Méditerranée. Nous avons étudié le sar commun (Diplodus sargus), poisson hermaphrodite protandre, entre une aire protégée et non protégée libre de toutes activités humaines. Nous avons observé une différence significative dans la structure démographique entre ces deux sites. Les plus jeunes femelles ont été capturées à l’extérieur de l’aire protégée. Ceci suggère une plasticité phénotypique dans l’âge d’inversion du sexe dans le cas d’une population exploitée. La différenciation génétique (FST = 0,007, p = 0,04) repose sur un seul locus et semble donc non fondée. Cela suggère des flux géniques importants ou une séparation trop récente des deux populations pour faire apparaître une différenciation génétique. Nos résultats sur le sar commun montrent que les poissons dans la réserve et à l’extérieur sont génétiquement très similaires, ce qui signifie que l’effet « réserve » mis en évidence semble réellement un effet au niveau démographique, et non le résultat de différences génétiques. Pour citer cet article : P. Lenfant, C. R. Biologies 326 (2003). 2003 Académie des sciences. Published by Éditions scientifiques et médicales Elsevier SAS. All rights reserved. Keywords: reserve effect; demographic structure; genetic differentiation; white sea bream; Diplodus sargusMots-clés : effet réserve ; structure démographique ; différenciation génétique ; sar commun ; Diplodus sargusE-mail address: lenfant@univ-perp.fr (P. Lenfant).
1631-0691/$ – see front matter 2003 Académie des sciences. Published by Éditions scientifiques et médicales Elsevier SAS. All rightsreserved. doi:10.1016/S1631-0691(03)00168-9
P. Lenfant / C. R. Biologies 326 (2003) 751–760Version française abrégée
d’autorisation. Le site choisi dans la réserve (PPA) estsitué à proximité d’un cap, sur des fonds rocheux de 20
Les réserves marines ont fait l’objet de nombreuses
à 30 m de profondeur. Le second site d’étude est situé
études, basées sur la comparaison de la biomasse et de
à l’extérieur de la réserve (UPA). Il a été sélectionné
la densité d’espèces cibles entre des zones protégées et
dans une zone (cap Béar), dont les caractéristiques
non protégées. Les poissons, de par leur intérêt com-
physiques sont proches de celles du site PPA et où la
mercial, ont souvent fait l’objet de ce type d’étude à
pression de pêche est importante. Les pêcheurs pro-
l’aide de méthodes non destructrices, comme le comp-
fessionnels utilisent les mêmes techniques que celles
tage visuel. Les sparidés sont très répandus dans la
employées dans la PPA. La pêche à la ligne amateur
Méditerranée nord-occidentale et constituent une res-
et la chasse sous-marine sont des activités courantes
source importante d’un point de vue commercial pour
sur cette côte rocheuse. Depuis plusieurs années, un
les pêcheries côtières. Le sar commun constitue, avec
championnat national de chasse sous-marine est or-
le sar à tête noire, Diplodus vulgaris (Saint-Hilaire,
ganisé chaque année sur cette zone. Les deux sites
1817) une des espèces les plus abondantes de l’infralit-
d’étude sont séparés d’environ 5 km. Dans le cadre de
toral rocheux et de l’herbier de posidonies. Il fait partie
notre échantillonnage, tous les poissons ont été captu-
du groupe des poissons nectobenthiques mésophages,
rés de nuit au fusil harpon, entre février et mai 1996.
caractérisés par des déplacements verticaux réduits et
Le temps de plongée et la profondeur ont été standar-
un potentiel de mouvements horizontaux important.
disés dans les deux sites. Chaque poisson a été me-
Les adultes sont hermaphrodites protandres. Les in-
suré (longueur standard), sexé puis disséqué pour ex-
dividus sont mâles à leur première maturité sexuelle
traire les otolithes (sagitta), le foie et un morceau de
(environ 3 ans) et deviennent femelles vers l’âge de
muscle. Les otolithes ont été inclus dans la résine,
5 ans. La reproduction a lieu au printemps, avec un
poncés et colorés au bleu de toluidine pour visualiser
maximum en avril sur les côtes catalanes françaises.
les stries annuelles. La validation de l’estimation de
Les zones de pontes ne sont pas précisément connues
l’âge a été réalisée par deux méthodes (évolution de
et semblent être différentes de l’habitat adulte. Après
l’accroissement marginal et injection de tétracycline).
un mois de durée de vie larvaire pélagique, les larves
Les échantillons ont été analysés par électrophorèse
recrutent sur des petits fonds, où elles y restent plu-
sur gels d’amidon horizontaux et gels d’acétate. Les
sieurs mois. Au-delà de 5 cm de longueur, les juvé-
fréquences alléliques et les F-statistics (FIS et FST)
niles quittent les zones de nurseries pour intégrer la
ont été calculés à partir du logiciel GENETIX 4.0.
population adulte. L’objectif de cette étude était de dé-
La « significativité » de l’écart à l’équilibre de Hardy–
crire les structures démographiques et génétiques des
Weinberg et de la différenciation génétique a été éva-
populations de sar commun et d’évaluer l’impact de la
luée par les tests exacts de Fisher (GENEPOP 3.1d).
protection sur les populations de la réserve marine de
La longueur standard des individus varie de 13,1 à
Cerbère–Banyuls, située sur la côte catalane française
29,2 cm dans le site non protégé (UPA : 18,4 − SE =
3,7 − n = 61) et de 15,3 à 27,0 cm dans la réserve
Les échantillons de sars communs ont été collectés
(PPA : 19,6 − SE = 2,6 − n = 70). La distribution des
dans deux sites des côtes méditerranéennes françaises
âges montre que les poissons sont nés entre 1988 et
proches de l’Espagne. Le premier site est localisé dans
1993 (respectivement des cohortes de huit à trois ans).
la réserve marine de Cerbère–Banyuls (650 ha), créée
Les distributions de la longueur standard et de l’âge
en 1974, et qui est constituée de deux zones : une zone
sont unimodales dans les deux sites. Les individus
partiellement protégée (585 ha) (PPA) et une zone to-
sont significativement plus grand et plus vieux dans la
talement protégée (65 ha) (TPA). La législation dans
réserve (tests de Mann–Whitney, longueur standard :
la réserve autorise la pêche récréative à la ligne, mais
Z = −3,51 − p < 0,001 ; âge : Z = −4,39 − p <
réglemente l’activité de pêche artisanale (cinq à huit
0,001). De plus, les femelles sont plus nombreuses
autorisations par an). La chasse sous-marine est stric-
que les mâles dans les deux sites, mais la distribution
tement interdite. Dans la TPA, toutes les activités sont
des femelles en fonction de l’âge est différente au sein
interdites, à l’exception des plongées scientifiques.
et à l’extérieur de la réserve (Pearson χ 2 ; femelles :
Aucun poisson n’a été capturé dans cette zone, faute
χ 2 = 21,37 − p = 0,001 ; mâles : χ2 = 1,16 − p =
P. Lenfant / C. R. Biologies 326 (2003) 751–760
0,88). Le mode de distribution des femelles est centré
sur les oursins, il n’est pas possible de valider l’une ou
sur les cohortes de quatre (1992) et six ans (1990),
respectivement pour les sites UPA et PPA. La cohorte
La présence d’individus plus grands et plus vieux
4 ans, qui représente plus de 30% dans l’UPA, est
dans les réserves s’explique par la réduction de l’ef-
quasiment absente dans la réserve (PPA). Inversement,
fort de pêche. Ces observations ont été signalées chez
les cohortes 6 ans et 7 ans sont proportionnellement
d’autres espèces de poissons et d’invertébrés. Dans
plus faibles à l’extérieur de la réserve. Les analyses
notre étude, la distribution des tailles individuelles est
génétiques ont porté sur 26 locus analysés dont 16 sont
positivement corrélée avec le degré de protection. Le
apparus polymorphes à 99% (13 locus polymorphes à
modèle classique décrit l’influence des réserves ma-
95%). Les principaux paramètres étudiés montrent de
rines sur les paramètres démographiques des popu-
faibles variations entre les deux sites. Aucun écart à
lations. Ces changements démographiques ont pour
l’équilibre de Hardy–Weinberg n’a été observé dans
conséquences, d’une part, une augmentation de l’âge à
les deux sites. La valeur de FST multilocus (0,007 −
la maturité sexuelle dans la réserve et, d’autre part, une
p = 0,04) est significative et est essentiellement due
augmentation de l’exportation (adultes et/ou progéni-
à un seul locus (GDA* : FST = 0,054 − p = 0,0003).
tures vers l’extérieur de la réserve). Nous n’avons pas
Tous les autres locus génèrent des valeurs de FST non
observé de modification de l’âge à la première matu-
rité sexuelle. En revanche, l’âge d’inversion sexuelle
La structure démographique et la distribution du
de mâle à femelle, qui se situe aux alentour de cinq
sexe en fonction de l’âge chez le sar commun sont
à six ans chez le sar commun, est modifié à l’exté-
affectées par la protection offerte par la réserve ma-
rieur de la réserve, puisque nous observons une forte
rine. La structure génétique des individus des deux
proportion de femelles de quatre ans. Ces résultats
sites montre une différenciation génétique induite par
complètent le modèle proposé classiquement dans la
un seul locus. Cependant, ceci ne semble pas suffi-
littérature. Le fait qu’un signal au niveau démogra-
sant pour conclure à une forte différenciation entre
phique soit détecté est en faveur d’un échange ré-
ces deux sites. Des études précédentes, réalisées dans
duit d’individus entre la réserve et l’extérieur. Néan-
cette même réserve, ont montré des résultats contra-
moins, à ce stade, nous ne pouvons pas conclure à une
dictoires. La première étude, réalisée en 1980 par
absence complète d’échange d’œufs et de larves par
comptage visuel, a montré que les poissons étaient
les courants. Pour tester cette hypothèse, nous avons
plus gros et plus nombreux dans la réserve (TPA) qu’à
utilisé les marqueurs génétiques. La structure géné-
l’extérieur (cap Béar). Cet effet serait dû principale-ment à une interdiction de la chasse sous-marine, mise
tique est différente entre les individus de la réserve
en place dès la création de la réserve en 1974. Dix
et ceux de l’extérieur. Cette différence n’est suppor-
ans plus tard, une seconde étude a été réalisée sur les
tée que par un seul locus (GDA*), ce qui limite notre
mêmes sites, avec des résultats parfois inverses. Deux
confiance dans cette différenciation. Dans notre étude,
hypothèses peuvent expliquer ces résultats contradic-
nous avons volontairement pris un nombre de locus
toires. (i) La première fait intervenir une dynamique
polymorphes conséquent (16 locus) pour nous rappro-
complexe, qui serait différente de celle classiquement
cher de la sensibilité des marqueurs moléculaires. Si
proposée, qui prédit que les individus âgés ou de
nous considérons que les marqueurs allozymiques sont
grande taille sont plus nombreux dans la réserve. Des
fiables, la similitude génétique que nous observons in-
études comparant l’abondance et la densité de popula-
dique un flux génique (migration d’adultes et/ou ex-
tions d’oursins à plusieurs années d’intervalle aboutis-
portation d’œufs et de larves par les courants) suffisant
sent également à ce type de résultats contradictoires.
pour maintenir les mêmes fréquences alléliques entre
(ii) La seconde envisage un biais dans l’échantillon-
les deux sites. Alternativement, si le flux génique est
nage. L’estimation par comptage visuel dépend du
réellement réduit, la similitude génétique peut résulter
comportement de chaque espèce, alors que la capture
d’une séparation récente des deux sites. Néanmoins,
des poissons de nuit permet un effort d’échantillon-
la première hypothèse semble la plus parcimonieuse,
nage indépendant du comportement. Sans étude com-
car le sar commun présente de fortes capacités de dis-
parative et au vu des résultats contradictoires obtenus
persion par la phase larvaire. L’effet « réserve » mis en
P. Lenfant / C. R. Biologies 326 (2003) 751–760
évidence semble donc réellement un effet au niveau
importance, no quantitative information is available
on the dynamics of sparid fish populations on rocky
La compréhension de l’effet « réserve » nécessite
shores, except for a few studies on the microhabitat
une bonne connaissance des communautés de pois-
requirements at settlement [12,13]. Adult fishes are
sons, mais pas uniquement en termes de densité et de
protandrous hermaphrodites. Fishes are male at the
biomasse obtenues par comptage visuel. Les présents
first sexual maturity (three years old) and then become
résultats suggèrent qu’il faut tenir compte des parti-
female at five years old [14]. The reproduction occurs
cularités du cycle biologique de chaque espèce cible,
during spring (maximum spawning in April on the
à savoir l’âge, le changement de sexe, etc. D’autres
French Catalan coasts) in areas that are not precisely
facteurs pouvant influencer la dynamique des espèces
known. The increase of capture by fishermen (trawl-
cibles, comme la courantologie et la pression de pêche,
ing) seems to indicate that fishes leave the rocky coasts
sont à prendre en considération, mais ne sont pas tou-
to migrate offshore to form several spawning groups.
jours disponibles dans toutes les aires marines pro-
After one month of pelagic larval drift [15], post-
tégées. Ces résultats confirment le potentiel des ré-
larvae settle in very shallow benthic areas, where they
serves marines en tant qu’outil de gestion des espèces
remain for several months [13]. After reaching 4.5 to
sensibles à la « surpêche ». En association avec les
5.5 cm in length, juveniles disperse outside the nursery
méthodes non destructrices, de nouvelles approches,
areas and integrate into the adult population [16].
telles que les marqueurs acoustiques, pourraient ac-
The aim of the present study was to describe the
croître nos connaissances sur l’effet « réserve » et les
demographic and genetic structures of Diplodus sar-
mécanismes d’exportation d’adultes. gus populations and evaluate the impact of protectionon populations living inside and outside the marine re-serve of Cerbère–Banyuls, located in French Catalo-
1. Introduction
Many studies have highlighted the effects of marine
reserves by comparing target species biomass and den-
2. Material and methods
sity within and outside a reserve. These studies haveinvolved fish populations because of their commercial
Samples of white sea bream were collected in two
interest and are based on visual census methods [1–
sites on the French Mediterranean coasts near Spain.
4]. With respect to invertebrate populations, studies
The first site is located in the Cerbère–Banyuls ma-
have focused on echinoids and bivalves [5–7], and gas-
rine reserve (650 ha) created in 1974, with two zones:
tropods [8,9]. To date, the ‘reserve effect’ on the white
a Partially Protected Area (PPA) (585 ha) and a To-
sea bream, Diplodus sargus (Linnaeus, 1758), was in-
tally Protected Area (TPA) (65 ha, created in 1979).
vestigated in 1980 and 1994–1995 and has become a
The second site in an unprotected area (UPA). These
model for studies of benthic fishes on rocky coasts [10,
two sites are separated by 5 km and provide two lev-
els of protection for the fish populations. The site se-
Sea bream fishes (family Sparidae) are widespread
lected in the PPA is located near a rocky cap with the
in the northwestern Mediterranean Sea and constitute
following physical characteristics: rocky bottom, 20–
an important fishery resource along its coasts for their
30 m deep. The marine reserve legislation allows ama-
high value. The white sea bream, Diplodus sargus
teur angling, but limits the commercial fishing activity
(Linnaeus, 1758), constitutes with Diplodus vulgaris
(gill nets) (5–8 accreditations per year). Spear fishing
(Saint-Hilaire, 1817) one of the most important fishes
is prohibited. In the TPA, all activities are prohibited,
of infra-littoral rocky bottoms and Posidonia ocean-
but scientific study. No fish was caught in this zone. ica beds (Linnaeus) Delile. It belongs to the group
The UPA site has been selected in an area (Cape Béar)
of necto-benthic mesophage fishes, which are char-
with equivalent physical characteristics than PPA site
acterised by a home range with low vertical move-
and subject to fishing pressure outside the reserve.
ment (from shallow water down to >50 m deep), but
There are no fishing restrictions at the UPA site. Pro-
high-potential horizontal movement [2]. Despite their
fessional fishermen (more than 10 fishing boats) use
P. Lenfant / C. R. Biologies 326 (2003) 751–760
the same gear as in the PPA (gill nets). Amateur an-gling and spear fishing are common activities on thisrocky coast. A national championship of spear fishingis organized every year (about 100 participants in fewkilometres) in the UPA site. Bell [1] ever mentionedthe same level of intense recreational fishing activitywhen he studied the fish communities in the Cerbère–Banyuls marine reserve.
All fishes were collected by spear fishing at night
between February and May 1996. Time and depthof dives were standardized in both sites. Each fishwas measured to the nearest millimetre (standardlength) and then dissected in order to remove otoliths(sagitta), the liver and a piece of muscle (1 or 2 g)and determine the sex. Otoliths were embedded inresin and sanded with sandpaper. The otoliths werecoloured with toluidine blue to visualise annual rings. The validation of the age determination was car-ried out using two methods: marginal increment andtetracycline injection [17]. For genetic analysis, tis-sues (liver and muscle) were stored in liquid ni-trogen (−196 ◦C) until electrophoresis. Each pieceof tissue was homogenized in an equal volume ofTris/EDTA/NADP buffer (pH 6.8). The homogenatewas centrifuged at 15 000 g for 30 min at 4 ◦C.
Fig. 1. Abundance (%) as a function of size class (standard length in
The supernatant was stored at −80 ◦C. The sam-
cm) and age of Diplodus sargus populations in an unprotected area
ples were then processed by routine electrophore-
(UPA) and a partially protected area (PPA).
sis on horizontal starch [17–19] and cellulose ac-etate plates (Titan III, Helena Laboratory). Twenty-six loci were analysed (muscle: CK*, GAPD-2*,
18.4, SE = 3.7, n = 61) and from 15.3 to 27.0 cm in
PT-1*, PT-2*, PT-3*, HPD-2*, GPI-2*, PGM*,
partially protected area (PPA: mean = 19.6, SE = 2.6,
EST-1*, AAT-1*, LDH-1*, ME*, αGPD*; liver:
n = 70). The age distribution showed that individuals
ADH*, GAPD-1*, LDH-2*, SDH*, ADA*, MDH*,
were born between 1988 and 1993, corresponding to
6PGD*, EST-3*, AAT-2*, GDA*, IDH*, SOD*, XO*)
cohorts 8+ and 3+, respectively. The standard length
with 16 polymorphic loci (99%). Allele frequencies
and age frequency distributions are unimodal in both
and F -statistics (FIS and FST estimated by Weir and
sites (Fig. 1) and individuals are significantly larger
Cockerham, [20]) were computed with GENETIX 4.0
and older in the PPA (Mann–Whitney tests, standard
software. The significance of deviations from Hardy–
length: Z = −3.51, p < 0.001; age: Z = −4.39,
Weinberg equilibrium and of genetic differentiation
p < 0.001). Additionally, females are more numerous
was assessed by Fisher’s exact test (GENEPOP 3.1d
than males in both sites but the frequency distribution
as a function of age is different for females withinand outside the marine reserve (Pearson χ 2; female:χ 2 = 21.37 − p = 0.001; male: χ2 = 1.16 − p =
3. Results
0.88) (Fig. 2). The mode of female distribution is the
cohort 4+ (1992) and 6+ (1990) for the UPA site andthe PPA site, respectively. Moreover, the cohort 4+,
The standard length of individuals ranged from
which represents more than 30% of the females in
13.1 to 29.2 cm in the unprotected area (UPA: mean =
the UPA site, is almost non-existent in the PPA site
P. Lenfant / C. R. Biologies 326 (2003) 751–760
Fig. 2. Male and female abundance (%) as a function of size class (standard length in cm) and age of Diplodus sargus populations in anunprotected area (UPA) and a partially protected area (PPA).
(less than 2%). Inversely, the cohorts 6+ and 7+ are
p = 0.0003). All others loci generate an FST value not
proportionally lower in the UPA site. 4. Discussion
Electrophoresis performed on 22 enzymes gave
a total of 26 loci with 16 and 13 loci showing
The comparison of standard length frequencies, sex
allelic variation at 99 and 95%, respectively (Table 1).
distribution with age and genetic structure of white sea
Principal genetic parameters (frequency of the most
bream populations in protected and unprotected areas
common allele, observed heterozygosity per locus and
shows that the demographic structure and the sex dis-
multilocus, polymorphism (95%) and mean number
tribution of D. sargus are affected by the protection af-
of alleles) showed only small variation between the
forded by the marine reserve. The genetic structure of
two sites (Table 1). No significant deviation from the
both sites shows a differentiation induced by only one
Hardy–Weinberg equilibrium was observed for both
locus (GDA*). This is not sufficient to conclude that
sites (UPA: FIS = 0.069, p = 0.96; PPA: FIS = 0.083,
strong differentiation exists between these two sites. p = 0.68). The FST(0.007, p = 0.04) was significant
Previous studies of fish populations in the marine
essentially due to only one locus (GDA*: FST = 0.054,
reserve of Cerbère–Banyuls, including the white sea
P. Lenfant / C. R. Biologies 326 (2003) 751–760
Table 1Frequency of the most common allele frequencies (100) of all polymorphic loci found in two sites of white sea bream, including enzyme codenumber (EC), and buffer and tissue with best resolution. Genetic variability data includes mean number of alleles, percentage of polymorphicloci (P0.05) and observed heterozygosity (Hobs) (TC 8.0: tris-citrate, pH = 8.0; TC 6.7: tris-citrate, pH = 6.7; TBE: tris-borate–EDTA ; Pasteuret al. [18])
bream, show contradictory results. Bell [1] conducted
and older individuals. Many studies comparing the
the first study between the totally protected area (TPA)
abundance and density of sea urchin populations show
and unprotected area (Cape Béar) by visual census
variable results. Sala and Zabala [24] monitored the
in 1980. This author showed that the density and the
abundance of Paracentrotus lividus within and outside
size of fish populations were significantly higher in the
the Medes Islands marine reserve (NE Spain) for three
marine reserve, especially for sparid fishes (Diplodus
years and reported a pattern of lower abundance in
spp.). The author considered that angling and netting
the reserve. Later, a similar survey did not found any
prohibitions alone would not induce a significant ef-
significance difference in density and mean size [25].
fect within 14 months between their introduction and
(ii) A bias in the sampling methods used in these stud-
his study. The increased density and larger size struc-
ies (visual census in Dufour et al., [22], spear fish-
ture of vulnerable fishes in TPA of the reserve may,
ing by divers at night in our study). Firstly, the visual
therefore, be largely due to the longer term prohibi-
census method depends on the behaviour of fishes in
tion of spear fishing. Ten years later, Dufour et al. [22]
relation to the diver. Avoidance phenomena such as
did not observe significant difference in density and
flight and hiding places can occur during the visual
size between the marine reserve and the same unpro-
census and bias the biomass results [26]. In contrast,
tected site (Cape Béar). The authors, in fact, some-
the method of spear fishing by night dives can be eas-
times observed an opposite effect (medium-sized indi-
ily standardized in terms of duration and depth. The
viduals more abundant outside the reserve in deep wa-
sampling effort is in this case independent of fish be-
ter). This absence of reserve effect may be explained
haviour, as fishes are sleeping and the diver inspects
by two hypotheses. (i) Complex population dynam-
each hole in the rocky substrate with a submarine light.
ics that are different from the process described by
Without a comparative analyse of these methods and
Sánchez Lizaso et al. [23] showing that the decrease in
considering contradictory results for sea urchin, we
fishing mortality should lead to a shift towards larger
cannot evaluate these alternative hypotheses. P. Lenfant / C. R. Biologies 326 (2003) 751–760
The presence of the larger and older individuals in
lation. This mechanism complements the model pro-
marine reserves is explained by the reduction of fish
posed by Sánchez Lizaso et al. [23]. The significant
mortality. A higher proportion of larger and older in-
signal in term of demographic structure in our study
dividuals in protected areas has already been found
indicates that the migration of adult fishes between the
for various fish species in the same Mediterranean
marine reserve and adjacent areas is restricted. Never-
reserve [1], and in other marine reserves [3,27–32].
theless, at this stage, we cannot conclude that there is
Similar results were found for invertebrates [5,7,8,31].
a complete lack of exchange by eggs and larvae using
Protected areas usually lead to increases in density,
currents. To test the hypothesis of a lack of exchange,
biomass, diversity and/or longevity relative to popula-
we used genetic markers (allozymes).
tions subject to fishing pressure [33,34]. In our study,
The genetic structure is different inside and out-
the distribution of individual size, thus the biomass
side the marine reserve, but this differentiation is sup-
and the longevity, are positively and significantly re-
ported by only one locus (GDA*), which limits our
lated to the protection level of D. sargus. Sánchez
confidence in the concluding genetic differentiation.
Lizaso et al. [23] described the possible changes in
In the context of the controversy of genetic method
population dynamics of exploited species in marine re-
(allozyme vs DNA), several authors suggest that the
serves. The decrease in fishing mortality induces an
polymorphism may be not as well detected using al-
increase in survival, which allows an increase in sev-
lozyme electrophoresis as using molecular markers
eral demographic factors: mean size, age, density and
such as RAPD or microsatellites. Allendorf and Seeb
biomass. These demographic changes have two poten-
[37] have tested for differences in the patterns of diver-
tial outcomes: (i) an increase in the age at maturity for
gence among different types of markers (allozymes,
fishes living in the marine reserve and (ii) an increase
nuclear DNA and mitochondrial DNA) in four spawn-
in the exportation of adults and/or offspring from
ing populations of sockeye salmon (Oncorhynchus
the marine reserve. We observed that females were
nerka). They found concordance among markers in the
smaller and younger outside the marine reserve than
amount of genetic variation within and among popula-
females caught inside the marine reserve (Fig. 2). But
tions. They concluded that it is important to examine
the white sea bream is a protandrous hermaphrodite
many loci when estimating genetic differentiation and
fish, which is male at the first maturity (three years old)
patterns of genetic exchange among populations. Re-
and becomes female at 5–6 years old. This situation is
cent work on fishes shows that allozymes may, in some
observed in the marine reserve, but outside, 4-year-old
cases, be more discriminating than molecular markers
females have the highest frequency. We did not ob-
[38]. In this study, we have voluntarily worked with
serve any increase of the age at maturity in the ma-
a large number of loci (16 polymorphic loci), which
rine reserve. Nevertheless, we can consider that fishes
allows detection of polymorphism equivalent to mole-
become female younger outside than inside the ma-
cular methods [39]. If we consider that allozymes are
rine reserve. It seems that the sexual inversion in fe-
reliable, then in the absence of exchange, both popula-
males presents a plasticity allowing individuals to be-
tions would have to evolve differently in terms of ge-
come female earlier outside than inside the marine re-
netic structure, essentially due to genetic drift and/or
serve. The mechanisms of sexual inversion have ever
different selective pressures within and outside the ma-
been studied in hermaphrodite fishes and show a high
rine reserve (fishing pressure, food abundance, demo-
complexity. In proterogyne species, the disappearance
graphic structure, etc.). In our case, the differentiation
of dominant males induces the sexual inversion of the
is minor, because it depends only on one locus. The
bigger female [35]. But this strategy is modified by
genetic similarity means that gene flow (adult migra-
the presence of other adjacent harems as in the an-
tion and/or exportation of eggs and larvae by currents)
gel fish, Centropyge ferrugatus (Randall and Burgess,
is sufficient to maintain the same allele frequencies be-
1972) [35,36]. In the case of hermaphrodite fishes, our
tween the marine reserve and the unprotected site lo-
data are the first to describe a decrease of age at sexual
cated about 5 km away. Alternatively, if the gene flow
inversion in an exploited fish population. The sex ratio
is restricted, the genetic similarity may result from the
could be regulated by density dependent phenomena
recent separation between both sites due to the recent
in relation to the proportion of each sex in the popu-
status of protection in the marine reserve, which was
P. Lenfant / C. R. Biologies 326 (2003) 751–760
created only 18 years ago. The divergence time may
their helpful comments on the first draft of the man-
have been too short to generate differences in allelic
uscript. E.B. Taylor helped in improving the English
frequency. Nevertheless, the first hypothesis seems to
be the most parsimonious one, because the white seabream has great dispersal capabilities during all itslife cycle (larval stage of one month, high mobility
References
of adult). Our data on the white sea bream show thatfishes inside and outside the marine reserve are very
[1] J.D. Bell, Effects of depth and marine reserve fishing restric-
similar genetically, which means that the ‘reserve ef-
tions on the structure of the rocky reef fish assemblage in theNorth-Western Mediterranean Sea, J. Appl. Ecol. 20 (1983)
fect’ is truly a demographic one, not the result of ge-
[2] J.-G. Harmelin, Structure and variability of the ichtyofauna in
a Mediterranean protected rocky area (National Park of Port-Cros, France), P.S.Z.N.I. Mar. Ecol. 8 (1987) 263–284.
[3] P. Francour, Pluriannual analysis of the reserve effect on
5. Conclusion
ichthyofauna in the Scandola natural reserve (Corsica, North-western Mediterranean), Oceanol. Acta 17 (1994) 309–317.
An understanding of the reserve effect requires the
[4] P. Sasal, E. Faliex, S. Morand, Population structure of Gobius
knowledge of fish communities not only in term of
bucchicchii in a Mediterranean marine reserve and in unpro-tected areas, J. Fish Biol. 49 (1996) 352–356.
density and biomass of target species that are obtained
[5] R.G. Cole, T.M. Ayling, R.G. Creese, Effects of marine reserve
by visual census. The present results suggest that we
protection at Goat Island, northern New Zealand, New Zeal. J.
have to consider the life cycle particularities of target
species, i.e. the age estimation, the variability of sex
[6] E. Sala, Fish predators and scavengers of the sea urchin
change or the recruitment not considered in this study. Paracentrotus lividus in protected areas of the north-westMediterranean Sea, Mar. Biol. 129 (1997) 273–283.
This information is not always available from visual
[7] C.F. Boudouresque, A. Caltagirone, J.R. Lefevre, V. Rico,
census and requires the catch of fishes. We are faced
R. Semroud, Macrozoobenthos de la réserve naturelle de
with the problem of the catch in a marine protected
Scandola (Corse, Méditerranée nord-occidentale), Analyse
area. Others factors, as currents, fishing pressures and,
pluriannuelle de l’« effet réserve », in : Economic impact of the
more generally, all pressures influencing directly or in-
Mediterranean coastal protected areas, Ajaccio, Medpan News,1991, pp. 15–20.
directly the target species are certainly important, but
[8] C.A. Moreno, K.M. Lunecke, M.I. Lepez, The response of an
are not available in each marine protected areas. The
intertidal Concholepas concholepas (Gastropoda) population
results confirm the potential of marine reserves as an
to protection from man in southern Chile and the effects on
effective mean of managing rocky reef species, espe-
benthic sessile assemblages, Oikos 46 (1986) 359–364.
[9] T.R. McClanahan, Kenyan coral reef-associated gastropod
cially those vulnerable to over-fishing. In combination
fauna: a comparison between protected and unprotected reefs,
with non-destructive methods (visual census by scuba
Mar. Ecol. Prog. Ser. 53 (1989) 11–20.
diving), new approaches such as acoustic tracking can
[10] E. Macpherson, F. Biagi, P. Francour, A. Garcia Rubies,
increase our knowledge of reserve effects [40]. This
J. Harmelin, M. Harmelin-Vivien, J.-Y. Jouvenel, S. Planes,
technique could allow us to understand the mechanism
L. Vigliola, L. Tunesi, Mortality of juvenile fishes of the genusDiplodus in protected and unprotected areas in the Western
of biomass export from the marine reserve with the
Mediterranean Sea, Mar. Ecol. Prog. Ser. 160 (1997) 135–147.
[11] S. Planes, J.-Y. Jouvenel, P. Lenfant, Density dependence in
post-recruitment processes of juvenile sparids in the littoral ofthe Mediterranean Sea, Oikos 83 (1998) 293–300.
[12] A. García-Rubies, E. Macpherson, Substrate use and temporal
Acknowledgements
pattern of recruitment in juvenile fishes of the Mediterraneanlittoral, Mar. Biol. 124 (1995) 35–42.
This study was financially supported by the Re-
[13] M. Harmelin-Vivien, J.-G. Harmelin, V. Leboulleux, Micro-
gional Council of Languedoc–Roussillon (France).
habitat requirements for settlement of juvenile sparid fishes onMediterranean rocky shores, Hydrobiologia 300–301 (1995)
I thank J.-L. Binche and A. Cazeilles (marine reserve
of Cerbère–Banyuls) for providing help and technical
[14] C. Martinez Pastor, M.L. Villegas Cuadros, Edad, crecimiento
assistance. I thank Drs S. Planes and R. Galzin for
y reproduccion de Diplodus sargus Linnaeus, 1758 (Spari-
P. Lenfant / C. R. Biologies 326 (2003) 751–760
dae) en aguas asturianas (norte de Espana), Bol. Inst. Esp.
[28] J.T. Bayles, A.A. Ramos-Espla, Some population parame-
ters as bioindicators for assessing the fish assemblage evo-
[15] L. Vigliola, M. Harmelin-Vivien, M.G. Meekan, Comparison
lution in a marine reserve, in: C.F. Boudouresque, M. Avon,
of techniques of back-calculation of growth and settlement
C. Pergent-Martini (Eds.), Qualité du milieu marin – Indica-
marks from the otoliths of three species of Diplodus from the
teurs biologiques et physico-chimiques, GIS Posidonie Publi-
Mediterranean Sea, Can. J. Fish. Aquat. Sci. 57 (2000) 1291–
cation, France, 1993, pp. 189–214.
[29] B.P. Ferreira, G.R. Russ, Population structure of the Leopard
[16] E. Macpherson, Ontogenetic shifts in habitats use and aggre-
coral grouper, Plectropomus leopardus, on fished and unfished
gation in juvenile sparid fishes, J. Exp. Mar. Biol. Ecol. 220
reefs off Townsville, Central Great Barrier Reef, Australia,
[17] P. Lenfant, Influence des paramètres démographiques sur la
[30] J.-G. Harmelin, F. Bachet, F. Garcia, Mediterranean marine
différenciation génétique intra- et inter-populations : le cas
reserves: fish indices as tests of protection efficiency, P.S.Z.N.I.
du poisson marin, Diplodus sargus (Linné, 1758), PhD thesis,
EPHE–University Paris-6, France, 1998.
[31] C. Pipitone, F. Badalamenti, G. D’Anna, B. Patti, Divi-
[18] N. Pasteur, G. Pasteur, F. Bonhomme, J. Catalan, J. Britton-
eto di pesca a strascio nel Golfo di Castellamare (Sicilia
Davidian, Manuel de génétique par électrophorèse des pro-
Nord-occidentale): alcune considerazione, Biol. Mar. Medit. 3
téines, collection Tech. & Doc., Lavoisier, Paris, 1987.
[19] P. Lenfant, S. Planes, Genetic differentiation of the white
[32] O. Reñones, J. Moranta, J. Coll, B. Morales-Nin, Rocky
sea bream, Diplodus sargus, within the Lion’s Gulf and the
bottom fish communities of Cabrera Archipelagos National
Ligurian Sea, J. Fish Biol. 49 (1996) 613–621.
Park (Mallorca, western Mediterranean), Sci. Mar. 61 (1997)
[20] B.S. Weir, C.C. Cockerham, Estimating F-statistics for the
analysis of population structure, Evolution 41 (1984) 1358–
[33] N.V. Polunin, C.M. Roberts, Greater biomass and value of
target coral reef fishes in two small Caribbean marine reserves,
[21] M. Raymond, F. Rousset, An exact test for population differ-
Mar. Ecol. Prog. Ser. 100 (1993) 167–176.
entiation, Evolution 49 (1995) 1280–1293.
[34] C.M. Roberts, Rapid build-up of fish biomass in a Caribbean
[22] V. Dufour, J.-Y. Jouvenel, R. Galzin, Study of reef fish
marine reserve, Cons. Biol. 9 (1995) 815–826.
assemblage. Comparison of population distributions between
[35] Y. Sakai, M. Kohda, Harem structure of the protogynous
depths in protected and unprotected areas over one decade,
angelfish, Centropyge ferrugatus (Pomacanthidae), Environ.
Aquat. Living Resour. 8 (1995) 17–25.
[23] J.L. Sánchez Lizaso, R. Goni, O. Renones, J.A. Garcia Char-
[36] Y. Sakai, Alternative spawning tactics of female angelfish
ton, R. Galzin, J.T. Bayle, P. Sanchez Jerez, A. Perez Ruzafa,
according to two different contexts of sex change, Behav.
A.A. Ramos, Density dependence in marine protected popula-
tions: a review, Environ. Conserv. 27 (2000) 144–158.
[37] F.W. Allendorf, L.W. Seeb, Concordance of genetic divergence
[24] E. Sala, M. Zabala, Fish predation and the structure of
among sockeye salmon populations at alozyme, nuclear DNA,
the sea urchin Paracentrotus lividus populations in the NW
and mitochondrial DNA markers, Evolution 54 (2000) 640–
Mediterranean, Mar. Ecol. Prog. Ser. 140 (1996) 71–81.
[25] E. Sala, M. Ribes, B. Hereu, M. Zabala, V. Alva, R. Coma,
[38] S. De Innocentiis, L. Sola, S. Cataudella, P. Bentzen, Al-
J. Garrabou, Temporal variability in abundance of the sea
lozyme and microsatellite loci provide discordant estimates
urchin Paracentrotus lividus and Arbacia lixula in the north-
of population differentiation in the endangered dusky grouper
western Mediterranean: comparison between a marine reserve
(Epinephelus marginatus) within the Mediterranean Sea, Mol.
and an unprotected area, Mar. Ecol. Prog. Ser. 168 (1998) 135–
[39] P.G. Parker, A.A. Snow, M.D. Shug, G.C. Booton, P.A. Fuerst,
[26] S. Jennings, N.V.C. Polunin, Biased underwater visual census
What molecules can tell us about populations: choosing and
biomass estimates for target-species in tropical reef fisheries,
using a molecular marker, Ecology 79 (1998) 361–382.
[27] A. García-Rubies, M. Zabala, Effects of total fishing pro-
M. Harmelin-Vivien, F. Badalamenti, L. Le Diréach, G. Ber-
hibition on the rocky fish assemblages of Medes Islands
nard (Eds.), Introduction guide of methods for selected eco-
marine reserve (NW Mediterranean), Sci. Mar. 54 (1990)
logical studies in marine reserves, GIS Posidonie publication,
Publication list of Werner Brannath A) Original papers in Statistics and Mathematics published in peer-reviewed journals Gutjahr G, Brannath W, Bauer P, (2010). A general approach to the conditional error rate principle with nuisance parameters. Biometrics, accepted. Brannath W and Bretz F (2010). Shortcuts for locally consonant closed test procedures. Journal of the American Statistical
L’ECHO VENDREDI 27 AVRIL 2012 David Brennan, directeur général d’AstraZeneca La crise? Pas pour tout le monde… d’Astra Zeneca jette l’éponge Après six ans à la tête du deuxième groupe pharmaceu- tique britannique, l’Américain doit céder sa place en raison de mauvais résultats financiers. «Superglue», JOHANN HARSCOËT, À LONDRES le chiffre d’aff




 Demographic and genetic structures of white sea bream
populations (Diplodus sargus, Linnaeus, 1758) inside and outside
Laboratoire d’ichtyoécologie tropicale et méditerranéenne, École pratique des hautes études, CNRS UMR 8046,
université de Perpignan, 52, av. de Villeneuve, 66860 Perpignan cedex, France
Received 6 September 2002; accepted 11 June 2003
Abstract
Demographic and genetic structures of white sea bream
populations (Diplodus sargus, Linnaeus, 1758) inside and outside
Laboratoire d’ichtyoécologie tropicale et méditerranéenne, École pratique des hautes études, CNRS UMR 8046,
université de Perpignan, 52, av. de Villeneuve, 66860 Perpignan cedex, France
Received 6 September 2002; accepted 11 June 2003
Abstract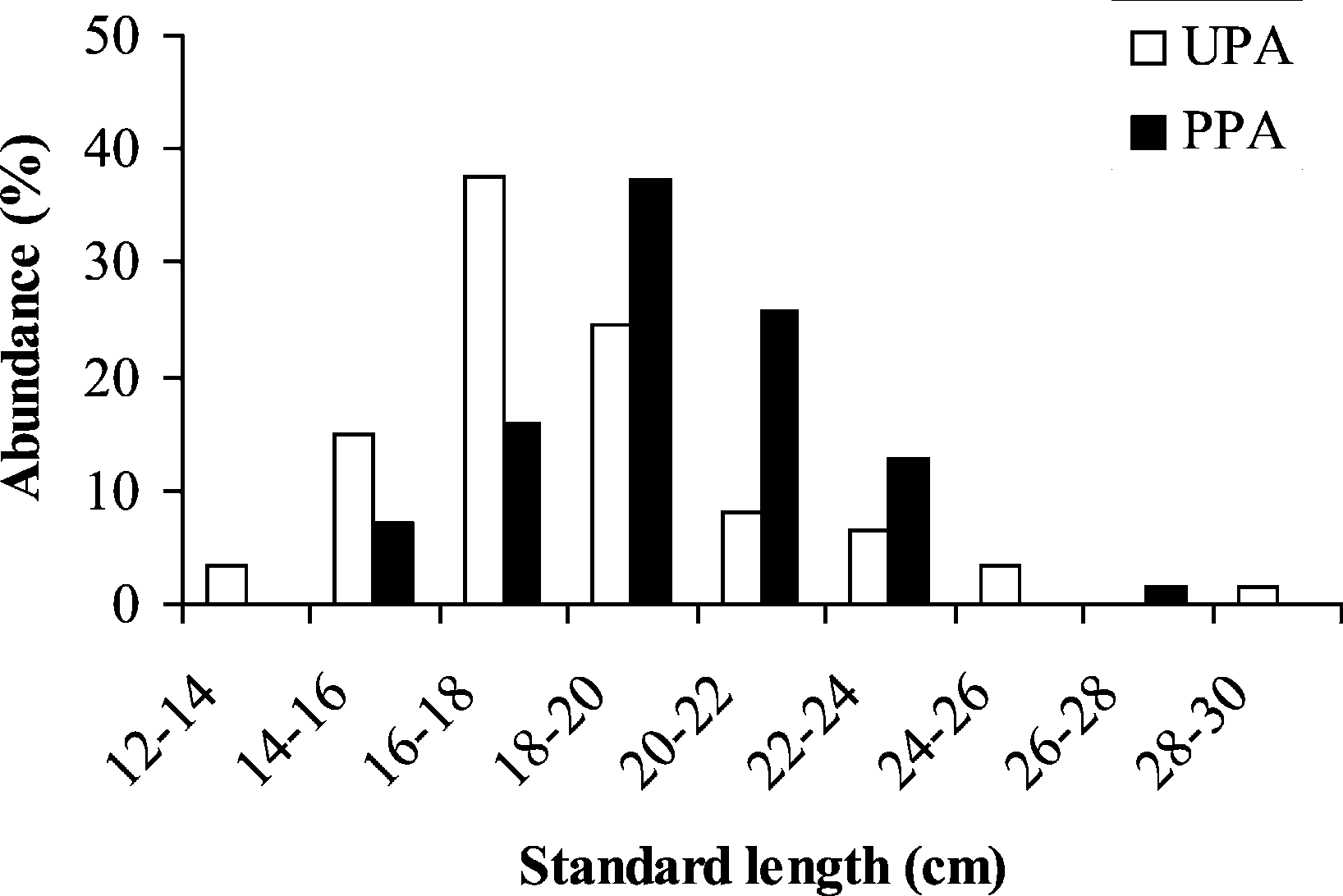
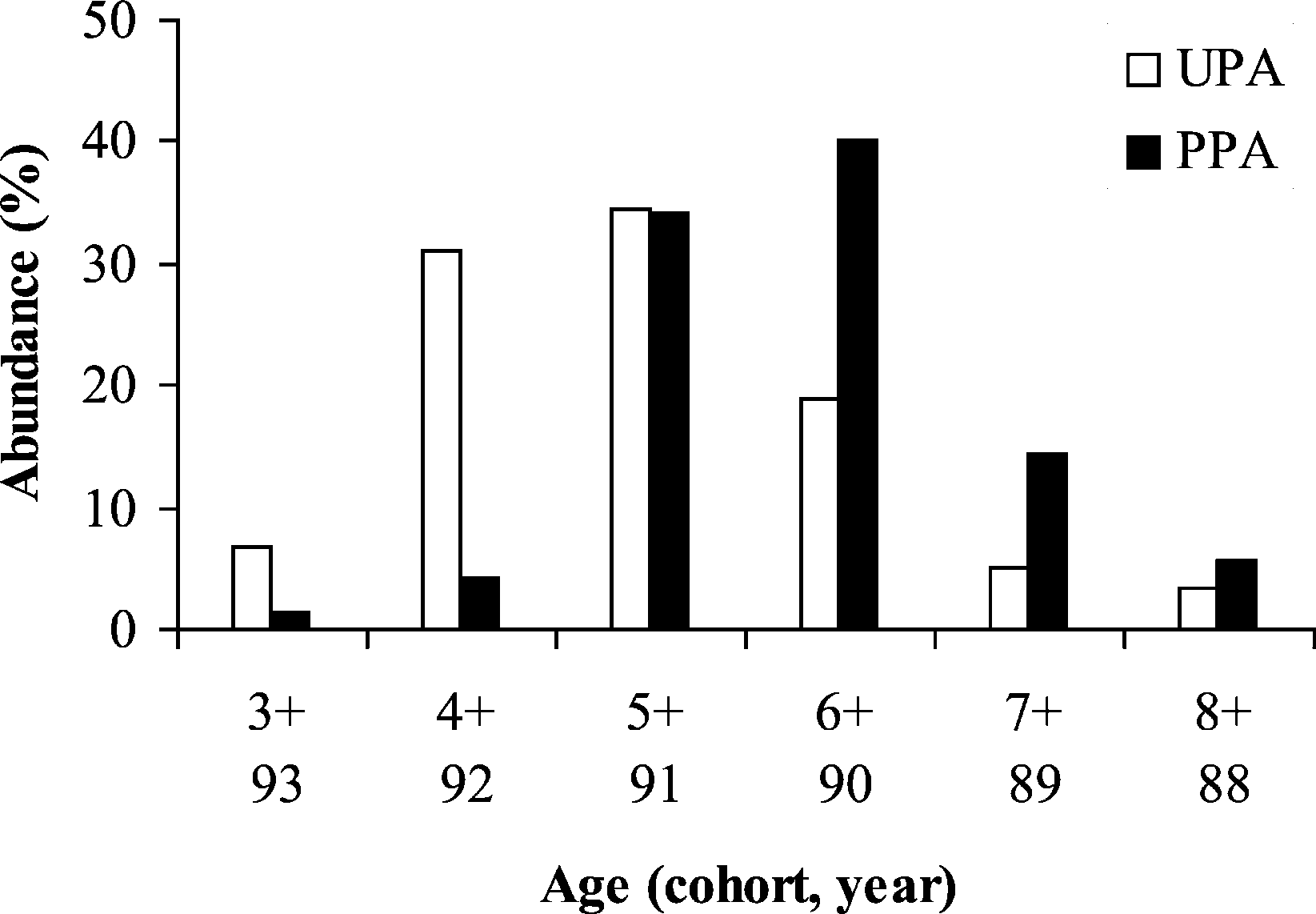 P. Lenfant / C. R. Biologies 326 (2003) 751–760
the same gear as in the PPA (gill nets). Amateur an-gling and spear fishing are common activities on thisrocky coast. A national championship of spear fishingis organized every year (about 100 participants in fewkilometres) in the UPA site. Bell [1] ever mentionedthe same level of intense recreational fishing activitywhen he studied the fish communities in the Cerbère–Banyuls marine reserve.
P. Lenfant / C. R. Biologies 326 (2003) 751–760
the same gear as in the PPA (gill nets). Amateur an-gling and spear fishing are common activities on thisrocky coast. A national championship of spear fishingis organized every year (about 100 participants in fewkilometres) in the UPA site. Bell [1] ever mentionedthe same level of intense recreational fishing activitywhen he studied the fish communities in the Cerbère–Banyuls marine reserve.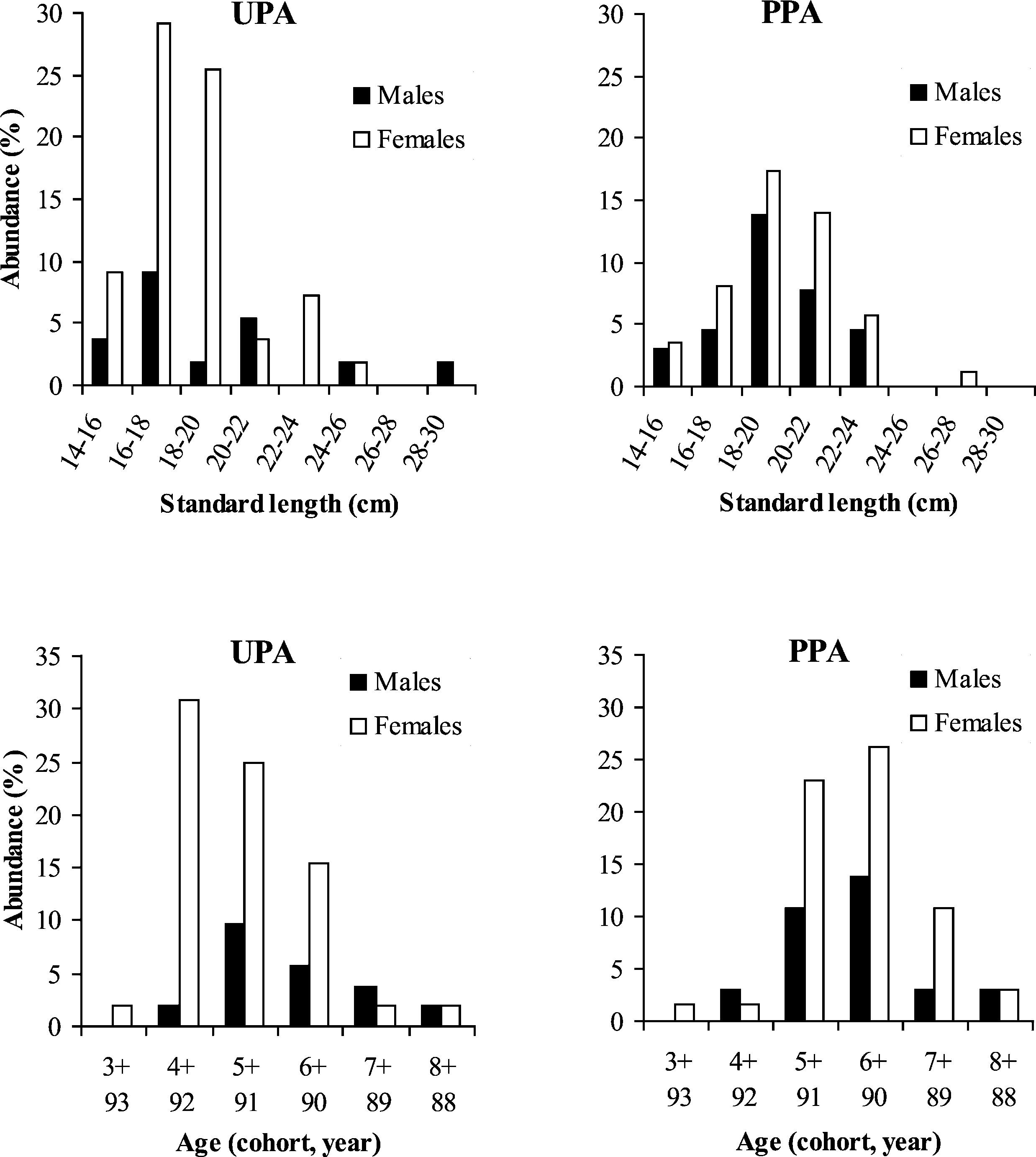 P. Lenfant / C. R. Biologies 326 (2003) 751–760
Fig. 2. Male and female abundance (%) as a function of size class (standard length in cm) and age of Diplodus sargus populations in anunprotected area (UPA) and a partially protected area (PPA).
P. Lenfant / C. R. Biologies 326 (2003) 751–760
Fig. 2. Male and female abundance (%) as a function of size class (standard length in cm) and age of Diplodus sargus populations in anunprotected area (UPA) and a partially protected area (PPA).



 P. Lenfant / C. R. Biologies 326 (2003) 751–760
Table 1Frequency of the most common allele frequencies (100) of all polymorphic loci found in two sites of white sea bream, including enzyme codenumber (EC), and buffer and tissue with best resolution. Genetic variability data includes mean number of alleles, percentage of polymorphicloci (P0.05) and observed heterozygosity (Hobs) (TC 8.0: tris-citrate, pH = 8.0; TC 6.7: tris-citrate, pH = 6.7; TBE: tris-borate–EDTA ; Pasteuret al. [18])
bream, show contradictory results. Bell [1] conducted
and older individuals. Many studies comparing the
the first study between the totally protected area (TPA)
abundance and density of sea urchin populations show
and unprotected area (Cape Béar) by visual census
variable results. Sala and Zabala [24] monitored the
in 1980. This author showed that the density and the
abundance of Paracentrotus lividus within and outside
size of fish populations were significantly higher in the
the Medes Islands marine reserve (NE Spain) for three
marine reserve, especially for sparid fishes (Diplodus
years and reported a pattern of lower abundance in
spp.). The author considered that angling and netting
the reserve. Later, a similar survey did not found any
prohibitions alone would not induce a significant ef-
significance difference in density and mean size [25].
P. Lenfant / C. R. Biologies 326 (2003) 751–760
Table 1Frequency of the most common allele frequencies (100) of all polymorphic loci found in two sites of white sea bream, including enzyme codenumber (EC), and buffer and tissue with best resolution. Genetic variability data includes mean number of alleles, percentage of polymorphicloci (P0.05) and observed heterozygosity (Hobs) (TC 8.0: tris-citrate, pH = 8.0; TC 6.7: tris-citrate, pH = 6.7; TBE: tris-borate–EDTA ; Pasteuret al. [18])
bream, show contradictory results. Bell [1] conducted
and older individuals. Many studies comparing the
the first study between the totally protected area (TPA)
abundance and density of sea urchin populations show
and unprotected area (Cape Béar) by visual census
variable results. Sala and Zabala [24] monitored the
in 1980. This author showed that the density and the
abundance of Paracentrotus lividus within and outside
size of fish populations were significantly higher in the
the Medes Islands marine reserve (NE Spain) for three
marine reserve, especially for sparid fishes (Diplodus
years and reported a pattern of lower abundance in
spp.). The author considered that angling and netting
the reserve. Later, a similar survey did not found any
prohibitions alone would not induce a significant ef-
significance difference in density and mean size [25].