Psychopharmacology (2000) 150:325–336Digital Object Identifier (DOI) 10.1007/s002130000442
Hongbo Zhu · Gordon A. Barr
Naltrexone-precipitated morphine withdrawal in infant ratis attenuated by acute administration of NOS inhibitorsbut not NMDA receptor antagonists
Received: 17 November 1999 / Accepted: 6 March 2000 / Published online: 19 April 2000 Springer-Verlag 2000
Abstract Rationale: There is increasing evidence that
the N-methyl-D-aspartate (NMDA) receptor and the ni-tric oxide system are involved in opiate dependence in
There is a large population of opiate-addicted childbear-
the adult rat, but whether these results in the adult apply
ing-aged women in the United States and each year un-
to the infant rat is unknown. Objectives: Here we exam-
known numbers of infants are born to these women
ined the effects of NMDA receptor antagonists and nitric
(Hutchings 1987). These human infants, born to mothers
oxide synthase (NOS) inhibitors, which reduce the opiate
who are exposed to opiate drugs such as heroin during
abstinence syndrome in adult animals, on morphine
their pregnancy, have a higher incidence of morbidity
withdrawal in the infant rat. Methods: Neonatal rats were
and mortality than do the offspring of nonaddicted wom-
injected with morphine sulfate (10.0 mg/kg) twice daily
en (Naeye et al. 1973; Connaughton et al. 1977; Wilson
for 6.5 days. On the 7th day, pups were injected with
et al. 1979). Although there is much research on opiate
NOS inhibitors (L-NAME or 7-NI), NMDA receptor an-
withdrawal in adult animals, there are few studies in the
tagonists (MK-801 or AP-5), or vehicle. After 15 min,
neonates (Barr and Jones 1994). Recent studies have
the pups were injected with naltrexone (1 mg/kg) to pre-
shown that when the behavioral repertoire appropriate to
cipitate withdrawal. Behavior for each pup was identi-
the age of the animal is examined, infant rat pups experi-
fied and recorded every 15 s for 10 min before naltrex-
ence opiate tolerance and dependence if the dams are ex-
one injection and 15 min after naltrexone injection.
posed to opiates during their pregnancy or the pups are
Results: Both L-NAME and 7-NI significantly reduced
treated directly with opiates (Jones and Barr 1995;
most withdrawal behaviors in the infant rat, a result in
Windh et al. 1995; Thornton and Smith 1997; Thornton
line with previous studies in the adult rat. In contrast,
AP-5 reduced some withdrawal behaviors but also in-
In spite of decades of research, the understanding of
creased others (e.g., moving paws). MK-801 was like-
the mechanisms underlying opiate tolerance, physical de-
wise ineffective in reducing most withdrawal behaviors
pendence, and withdrawal is still very limited (Trujillo
and increased certain withdrawal behaviors (walking and
1999). It has been suggested that multiple mechanisms op-
wall climbing). Conclusions: In the infant rat, the pro-
erate in these processes (Thorat et al. 1994). Recently,
duction of nitric oxide is involved in opiate withdrawal
there have been a number of reports from a variety
whereas the NMDA receptor may not yet be functionally
active or may play only a minor role.
(NMDA) receptor antagonists (both noncompetitive andcompetitive) (Akaoka and Aston-Jones 1991; Trujillo and
Key words Opiate withdrawal · Neonatal · 7-NI ·
Akil 1991a, 1991b) and nitric oxide synthase (NOS) in-
hibitors (Adams et al. 1993; Cappendijk et al. 1993;Kimes et al. 1993; Vaupel et al. 1995a, 1995b) can inhibitthe development and expression of mu opioid dependence
This study was supported by grants DA 06600 and DA 00325
in the adult rodent. It has been hypothesized that NMDA
from the National Institute on Drug Abuse (NIDA) to G.A.B.
receptor, its second messenger system, and the productionof nitric oxide (NO) via the activation of the NMDA re-
ceptor play pivotal roles (Elliott et al. 1995; Thorat et al.
Biopsychology Doctoral Program, Department of Psychology,Hunter College, City University of New York, 695 Park Avenue,
1994; Mao et al. 1995; Inturrisi 1997; Mayer and Mao
1999; Trujillo 1995, 1999; Vaupel et al. 1995a, 1995b).
It is not known whether these mechanisms in adults
G.A. BarrNew York State Psychiatric Institute, New York, NY 10032, USA
apply to infants. The neonatal CNS is both structurally
and functionally different from that of the adult, and sig-
Table 1 Behavioral definitions
nificant changes in opioid actions occur both prenatally
and postnatally (Barr 1992, 1993; Fitzgerald 1995). Atthe same time, the NMDA receptor, which is believed to
play an essential role in opiate dependence, undergoes
qualitative and quantitative changes during development
(Kalb et al. 1992; Hori and Kanda 1994; Kalb and Fox
Turning the body over at least one full rotation
1997). There are significant developmental alterations
Bodily contact with one or more littermates
both in the density of the receptor (Tremblay et al. 1988;
Morin et al. 1989; Represa et al. 1989) and in the sensi-
Placing at least two forepaws on the wall of the
tivity to magnesium (Ben-Ari et al. 1988; Bowe and
Nadler 1990; Morrisett et al. 1990) of the NMDA recep-tor during the course of development. Thus, the pharma-cological effects of NMDA antagonists and NOS inhibi-
Morphine treatment and behavior observation
tors in the infant may not be necessarily comparable tothose in the adult. In fact, there is at least one study (Bell
Pups were tattooed with India ink (Geller and Geller 1966), whichwas injected into one or two paws to label individual pups perma-
and Beglan 1995) reporting negative results of the ef-
nently in each litter. To induce morphine dependence, all pups in a
fects of an NMDA receptor antagonist (MK-801) on the
litter were removed from the dam and individual rats were inject-
development of tolerance to morphine in the neonatal
ed with morphine sulfate (IP, 10 mg/kg) twice daily (10 a.m. and
rat, which is in contrast to reports in the adult rat
6 p.m.) for 6.5 days. The last injection was in the morning of the
7th day. In the afternoon of the 7th day, animals were transportedfrom the animal facility to our laboratory in plastic tubs with wood
The present studies investigated the pharmacological
chip bedding and placed in an observation chamber maintained at
effects of acute administration of MK-801, a noncompet-
approximately 33°C. Pups were weighed and subjected to various
itive NMDA receptor antagonist (Trujillo and Akil
drug treatment procedures (detailed in experiment 1 and experi-
1991a), AP-5, a competitive NMDA antagonist (Akaoka
ment 2). The order of treatment conditions was assigned randomlywithin each experiment. After the drug treatment, the pup was
and Aston-Jones 1991), NG-nitro-L-arginine methyl ester
then placed back into the observation chamber with the remainder
(L-NAME), a nonselective NOS inhibitor (Kimes et al.
of the litter (without the dam). Fifteen minutes later, naltrexone
1993; Vaupel et al. 1995a, 1995b), and 7-nitroindazole
was injected to precipitate withdrawal. The behavior of the pups
(7-NI), a neural selective NOS inhibitor (Kimes et al.
was observed for 10 min before the injection of naltrexone (5 minafter the treatment with test drugs) and 15 min after the injection
1993; Vaupel et al. 1995a, 1995b), on the naltrexone-
of naltrexone. During these observation periods, the behavior of
precipitated morphine withdrawal in the 7-day-old rat
the pups was identified every 15 s and recorded on a checklist by
pup. Although these compounds were reported to inhibit
an observer (see Table 1 for definitions of behavior included in the
both the development and the expression of opiate de-
checklist). When the observation for a specific pup ended, the pup
pendence, or withdrawal in the adult (Herman et al.
was anesthetized and placed back into the litter to keep the littersize unchanged. The next pup was then tested until all treatment
1995), the present studies focused attention on their po-
groups were completed. Animals were sacrificed with sodium
tential in blocking various withdrawal signs in the neo-
pentobarbital at the conclusion of the experiments.
natal rat. It was our hypothesis that these drugs shouldattenuate opiate withdrawal behaviors in the infant rat,even though any direct and meaningful comparison with
the adult may be difficult since withdrawal behaviorsdisplayed by the infant rat are very different from those
of the adult rat (Jones and Barr 1995; Windh et al. 1995;Thornton et al. 1997).
As described, in the afternoon of the 7th day, morphine-dependent rat pups were transported from the animal fa-cility to our laboratory. The pups were treated with saline
(IP, n=16, control for MK-801 and L-NAME, or ICV,n=8, control for AP-5), dimethylsulfoxide (DMSO) (IP,
n=8, control for 7-NI), MK-801 (IP, 0.01, 0.05,
All animal procedures were in accordance with the “Principles of
0.25 mg/kg, n=8 for each dose), AP-5 (ICV, 3, 30,
laboratory animal care” (NIH publication, 1996). The subjects
300 nmol, n=8 for each dose except for 300 nmol, which
were the offspring of Long-Evans hooded rats bred in our labora-
was terminated after four pups), L-NAME (IP, 15, 75,
tory. Parent animals were housed in plastic tubs with wood chips
375 mg/kg, n=8 for each dose), or 7-NI (IP, 5, 15,
in a colony room maintained at 22–24°C on a 12-h light/12-h darkphotocycle with light onset at 7 a.m. The breeding colony existed
45 mg/kg, n=8 for each dose), respectively. For each spe-
in a separate room with minimal disturbances except for normal
cific litter (N=16), eight pups were used. MK-801 and L-
cleaning, feeding and record keeping. Parent animals had Purina
NAME, including the respective control groups, were
Lab Chow (Purina 5012) and water available ad libitum. Cages
tested concurrently in the same litter. AP5 and 7-NI, in-
were checked twice daily, at approximately 10 a.m. and 6 p.m.
cluding the respective control groups, were also tested
Pups found at either time were termed 0 days of age. After parturi-tion, litters were culled to ten pups without regard for the ratio of
concurrently in the same litter. All drugs were delivered
via IP injection except AP-5 (and its control group),
which was delivered via ICV injection (Carden et al.
one period, behavior occurrences were summed for the
1991). AP-5 is not effective in passing the blood-brain
10-min observation period. For the data collected in the
barrier, and comparable studies (Akaoka and Aston-
post-naltrexone period, the 15-min observation was di-
Jones 1991) used similar techniques to deliver this com-
vided into three different time periods, each consisting of
pound directly into the CNS. During the ICV injection,
5 min; behaviors were summed in each time period,
the pup was held with its head gently bent forward and
which was treated as a within-subjects variable. For both
down. A beveled 30-gauge needle attached to a Hamil-
pre-naltrexone and post-naltrexone periods, all doses of
ton syringe was introduced into the cisterna magna and
the same drug were injected within a single litter, and the
4 µl of the drug was injected over a 30-s period. The nee-
drug dose effect was treated as a within-subjects vari-
dle was left in position for another 30 s, then removed.
Control groups received the same volume of saline in asimilar manner. We did not deliberately match the num-ber of males and females in each treatment group since
previous studies (Jones and Barr 1995) on the opiatewithdrawal in the 7-day-old rat show that gender is not a
factor in opiate withdrawal in rats of this age. Fifteenminutes after pre-treatment, the pup was injected with
naltrexone (IP, 1.0 mg/kg) to precipitate withdrawal andreturned to the litter. Behavior of the pup was observed
Treatment with various doses of 7-NI and L-NAME did
for 10 min before the treatment of naltrexone (pre-
not significantly alter the behaviors of the morphine-de-
naltrexone period) and 15 min after naltrexone injection
pendent rat (Table 2 ). Except for some baseline activi-
(post-naltrexone period). Data collected from the pre-
ties, all experimental groups largely remained quiet.
naltrexone period were used to analyze the effects of thedrugs on the morphine-dependent rat without the influ-ence of naltrexone. Any potential order effect was mini-
mized since the doses were randomly assigned (Latinsquare design) and the observer was blind to doses and
The results for the post-naltrexone period show that
drugs. We did not include saline control groups for mor-
treatment with both 7-NI and L-NAME significantly re-
phine treatment because previous reports by our labora-
duced most of the morphine withdrawal behaviors pre-
tory (Jones and Barr 1995) and others (Windh et al.
cipitated by naltrexone in infant rats (Table 3 ). We ana-
1995; Thornton and Smith 1997; Thornton et al. 1997)
lyzed a total of three possible effects: dose effects, time
had clearly demonstrated that saline-treated rat pups of
effects and interaction between dose and time. Both 7-NI
comparable age basically remain quiet and do not dis-
and L-NAME significantly dose dependently decreased
head moves, moving paws, rolling and together behav-iors and increased quiet behavior. In addition, pre-treat-ment with 7-NI significantly decreased walking behavior
in the infant rat; pre-treatment with L-NAME had no sig-nificant effect on walking behavior. We failed to find any
Separate statistical analyses were conducted for the four
significant effect on wall-climbing behavior for pre-
drugs. For each behavior, a one-way analysis of variance
treatment with either 7-NI or L-NAME. The dose effects
(ANOVA) was conducted for the pre-naltrexone period
of 7-NI and L-NAME on naltrexone-precipitated mor-
and a two-way ANOVA was conducted for the post-nal-
phine withdrawal in the 7-day-old rat are shown in
trexone period. For the data collected in the pre-naltrex-
Table 2 Behavior occurrences after treatment of NOS inhibitors but before treatment of naltrexone in the morphine-dependent rat of 7 days old
Note: cell entries are mean occurrences of behavior ± SEM. There were no significant differences between any treatment condition andthe corresponding control group (N = 8 per cell)
Table 3 Results of analyses of variance for withdrawal behav-
iors in NOS inhibitor treated 7-day-old rats (bold type P<0.05)
significantly attenuated the following behaviors: headmoves, together and walking, but increased quiet behav-
ior and intensified moving paws behavior. The dose ef-fects of MK-801 and AP-5 on naltrexone-precipitated
Treatment with various doses of MK-801 and AP-5 did
morphine withdrawal in the 7-day-old rat are shown in
not significantly alter the behaviors of the morphine-
dependent rat (Table 4). Except for some baseline activi-ties, all experimental groups largely remained quiet. Thehighest dose of AP-5 (300 nmol) sedated all treated sub-
jects (a total of four rat pups; simple reflexes disap-peared in these subjects) and we terminated the test for
The results of experiment 1 suggest that neither of the
this specific dose and the data are not included here.
two NOS inhibitors we tested significantly altered theoccurrences of various behaviors in the 7-day-old rat pri-or to naltrexone injection, whereas both 7-NI and L-
NAME were very effective in depressing various with-drawal behaviors precipitated by naltrexone. Thus, it
The results for the post-naltrexone period show that
seems that NOS inhibitors selectively attenuated with-
treatment with either MK-801 or AP-5 had unexpected
drawal behaviors. Both 7-NI and L-NAME significantly
effects on the morphine withdrawal syndrome (Table 5).
decreased head moves, moving paws, rolling and togeth-
MK-801 significantly decreased head moves, moving
er behaviors and increased quiet behavior. In addition,
paws, together and quiet behaviors. At the same time,
pre-treatment of 7-NI also significantly decreased walk-
pre-treatment with MK-801 significantly increased walk-
ing behavior in the infant rats. Although we failed to find
ing and wall-climbing behaviors. Treatment with AP-5
any significant results of wall-climbing behavior for ei-
Fig. 1 Dose effect of 7-NI. Ordinate mean occurrences (mean ± Fig. 2 Dose effect of L-NAME. Ordinate mean occurrences (mean
SEM) in 5 min of various opiate withdrawal behaviors (for defini-
± SEM) in 5 min of various opiate withdrawal behaviors (for defi-
tion see Table 1 ) in the 7-day-old rat. Abscissa treatment, log
nition see Table 1 ) in the 7-day-old rat. Abscissa treatment, log
scale (units: mg/kg). Dose effects were significant for all with-
scale (units: mg/kg). Dose effects were significant for all with-
drawal behaviors except walking and wall climbing
Table 4 Behavior occurrences after treatment of NMDA antagonists but before treatment of naltrexone in the morphine-dependent rat 7 days old
Note: Cell entries are mean occurrences of behavior ± SEM. There were no significant differences between any treatment condition andthe corresponding control group (N=8 per cell)aThis dose of AP-5 sedated all treated subjects (a total of four rat pups; simple reflexes disappeared in these subjects) and the test forthis specific dose was terminated
Table 5 Results of analyses of variance for withdrawal behav-
iors in NMDA antagonist treat-ed 7-day-old rats (bold type
ther 7-NI or L-NAME and walking behavior for L-
crease for pups pre-treated with MK-801. MK-801 was
NAME, this was very likely due to the fact that walking
reported to increase locomotion activities in adult rats
and wall climbing are very rare withdrawal behaviors in
(“PCP like” effect) (Herman et al. 1995). It is possible
the infant rat (Jones and Barr 1995), and thus the statisti-
that the observed increase in walking and wall climbing
cal power for these behaviors is considerably lower than
in the 7-day-old rat was due to the “PCP like” effect of
for other withdrawal behaviors that occurred more fre-
MK-801 independent of withdrawal. There are several
quently. Our results on the effect of NOS inhibitors in at-
reasons why this is unlikely. First, MK-801 did not sig-
tenuating withdrawal are in line with the findings by oth-
nificantly alter the occurrence of walking and wall-
ers in adult rats (Kimes et al. 1993; Vaupel et al. 1995a,
climbing behavior in the 7-day-old rat which was not ex-
1995b) and mice (Thorat et al. 1994).
periencing opiate withdrawal (Table 4); instead, MK-801
The effects of the NMDA antagonists are more diffi-
only increased these behaviors after the naltrexone chal-
cult to interpret. On the one hand, both MK-801 and AP-
lenge. Therefore, it seems that MK-801 selectively in-
5 attenuated some of the withdrawal behaviors; on the
creased withdrawal behaviors in the 7-day-old rat. Sec-
other hand, both of them increased some. The effects of
ond, in an ongoing study we also failed to find any sig-
MK-801 were especially unexpected. MK-801 dose de-
nificantly increased activity elicited by comparable dos-
pendently intensified walking and wall climbing and re-
es of MK-801 alone, MK-801 with chronic morphine but
duced the quiet behavior. Because we did not record
not naltrexone, or MK-801 with naltrexone but not
head moves and moving paws behaviors if the rat pup
chronic morphine in the 7-day-old neonatal rat. Third,
was walking or wall climbing, for MK-801, the attenua-
MK-801 induced locomotion activities only at consider-
tion of head moves and moving paws is most likely due
ably higher doses (Trujillo 1995). Lastly, when MK-801
to the robust walking and wall-climbing behavior exhib-
was administered to preweanling rats (12- to 19-day-
ited in the rat pups pre-treated with various dose of MK-
old), the “PCP like” effects of increased locomotion
801. Thus, overall withdrawal behaviors appeared to in-
were much less robust in the younger pups even within
Fig. 3 Dose effect of MK-801. Ordinate mean occurrences (mean Fig. 4 Dose effect of AP-5. Ordinate mean occurrences (mean ±
± SEM) in 5 min of various opiate withdrawal behaviors (for defi-
SEM) in 5 min of various opiate withdrawal behaviors (for defini-
nition see Table 1 ) in the 7-day-old rat. Abscissa treatment, log
tion see Table 1 ) in the 7-day-old rat. Abscissa treatment, log
scale (units: mg/kg). Dose effects were significant for all with-
scale (units: nmol). Dose effects were significant for all withdraw-
drawal behaviors except rolling. Note ordinate scale changes com-
al behaviors except rolling and wall climbing
that age range (Scalzo and Burge 1994). Since we found
no increased locomotion or wall climbing in pups givenMK-801, and our subjects were much younger than the
youngest pups demonstrating a “PCP like” effect, it isnot probable that the increased walking and wall climb-
Morphine dependence was established in infant rats as
ing seen after MK-801 administration in pups undergo-
described in experiment 1, and in the afternoon of day 7
ing withdrawal is due to the MK-801 alone.
rats were injected with saline (IP, n=32) or MK-801 (IP,
Since we used only one dose of naltrexone to precipi-
0.05 mg/kg, n=32) as pre-treatment. Fifteen minutes la-
tate withdrawal in experiment 1, there was a possibility
ter, each pup from both the saline pre-treated group and
that the intensification of walking and wall-climbing be-
the MK-801 pre-treated group was injected with naltrex-
haviors was an interaction between the MK-801 and the
one (IP, 0.03, 0.1, 0.3 or 1 mg/kg, n=8 for each dose) to
test concentration of the naltrexone (1 mg/kg). There-
precipitate withdrawal. All of the doses were randomly
fore, we tested the interaction between MK-801 and
assigned and the observer was blind to each dose and
multiple doses of naltrexone in experiment 2.
drug. The pup that received the naltrexone injection wasthen returned to the litter and its behavior observed for atotal of 15 min. A total of eight pups from each litterwere injected and tested in this manner. Table 6 Results of the analysis of the interaction between MK- 801 and naltrexone (bold type P<0.05) (MK-801×naltrexone indi- cates the interaction between MK-801 and naltrexone) Fig. 5 Interaction between dose of naltrexone and pre-treatment of MK-801. Ordinate mean occurrences (mean ± SEM) in 15 min of various opiate withdrawal behaviors (for definition see Table 1) in the 7-day-old rat. Abscissa naltrexone dose, log scale (units:
A two-way ANOVA was conducted for each behavior,which reflected its occurrences for 15 min. Unlike in ex-
(Table 6). Treatment of MK-801 significantly decreased
periment 1, the time factor was not analyzed in experi-
head moves and quiet behaviors, and, at the same time,
ment 2. All doses of naltrexone were injected within a
significantly increased walking and wall-climbing be-
single litter, and the drug dose effect was treated as a
haviors. We failed to find any significant effect of pre-
within-subjects variable. The control group (four pups)
treatment with MK-801 for the following behaviors:
and the MK-801 group (four pups) were from the same
moving paws, rolling and together. For no behavior was
litter and the MK-801 treatment was treated as a within-
there a significant dose effect of naltrexone. There was a
significant interaction between MK-801 and naltrexonefor the together behavior only.
The effects of pre-treatment with MK-801 at different
doses of naltrexone on withdrawal behaviors in the in-fant rat are depicted in Fig. 5.
Compared to the control group (saline pre-treatment),pre-treatment with MK-801 had significant effects onmorphine withdrawal behaviors precipitated by naltrex-
one in infant rats. We analyzed three possible effects: theeffects of the pre-treatment with MK-801 vs saline, the
The results of experiment 2 are generally in agreement
dose effects of the naltrexone and the interaction be-
with the results on the effects of MK-801 in experiment 1.
tween naltrexone and pre-treatment with MK-801
The interaction between treatment of MK-801 and nal-
trexone only exists for the together behavior, which de-
ment period (2–4 days) (Vaupel et al. 1995a, 1995b). In
clined with the high dose of MK-801 in experiment 1.
our experiment 1, the rat pups had been treated with
Unlike experiment 1, we failed to find any significant ef-
morphine considerably longer (6.5 days) before NOS in-
fect of pre-treatment of MK-801 on the moving paws be-
hibitor pre-treatment and naltrexone challenge, and we
havior. This difference may be due to the fact that, in ex-
did not find any increased withdrawal signs for either 7-
periment 2, we adopted a medium dose of MK-801
NI or L-NAME. Although we failed to find any signifi-
(0.05 mg/kg) and the dose effect of MK-801 in decreas-
cant effects on walking and wall-climbing behaviors for
ing moving paws behavior exhibited in experiment 1 was
L-NAME and on wall-climbing behavior for 7-NI, as ex-
significant only for the highest dose (0.25 mg/kg). As in
plained earlier, it is likely because these are rarely occur-
experiment 1, regardless of the dose of naltrexone, MK-
ring behaviors. Though statistically nonsignificant, the
801 decreased head moves and quiet behavior whereas it
mean occurrences of both wall-climbing behavior for the
increased the occurrences of walking and wall climbing.
7-NI pre-treatment group and walking behavior for the
MK-801 did not have any significant effect on the roll-
L-NAME pre-treatment group dose dependently de-
ing behavior. For the same reasons described in experi-
ment 1, we believe that the reduced head moves is likely
Vaupel et al. compared the opiate withdrawal attenua-
to be confounded by increased walking and wall climb-
tion effect of 7-NI with other NOS inhibitors (L-NNA,
ing and cannot be regarded as a sign of attenuation of
L-NAME, L-NIO) and found that 7-NI attenuated more
withdrawal for MK-801. Together with the results from
signs of opiate withdrawal than did L-NNA, L-NAME or
experiment 1, the results of experiment 2 confirmed that
L-NIO in the adult rat (Vaupel et al. 1995a). The results
MK-801 is ineffective in suppressing the naltrexone-pre-
of experiment 1 extend their data to the neonatal rat. All
cipitated morphine withdrawal in the neonatal rat.
withdrawal behaviors suppressed by L-NAME were alsoattenuated by 7-NI, and walking behavior was inhibitedby 7-NI but not L-NAME. Unlike other NOS inhibitors,
7-NI selectively inhibits neuronal NOS (Moore et al. 1993; Vaupel et al. 1995a, 1995b) and, therefore, it lacks
There are two conclusions that can be drawn from these
the capacity to elevate blood pressure in both untreated
experiments. First, acute administration of NOS inhibi-
and morphine-dependent rats (Vaupel et al. 1995a,
tors is quite effective in attenuating naltrexone-precipi-
1995b). The implication of these studies is that com-
tated morphine withdrawal in 7-day-old rat pups. Sec-
pounds like 7-NI should rank high on the list of drugs
ond, acute administration of NMDA antagonists is not
that have potential as a treatment for alleviating clinical
effective in inhibiting most of the naltrexone-precipitated
signs of opiate withdrawal in human infants.
morphine withdrawal behaviors in 7-day-old rat pups.
Since Trujillo and colleagues (Trujillo and Akil 1991a)
In the adult rat, L-NAME, 7-NI and other NOS inhibitors
first reported that the noncompetitive NMDA antagonist
have been widely reported to reduce opiate withdrawal
MK-801 inhibits the development of morphine tolerance
symptoms such as wet dog shakes, diarrhea, and groom-
and dependence in the adult rat, their findings have been
ing (Kimes et al. 1993; Vaupel et al. 1995a, 1995b).
widely replicated (Marek et al. 1991a, 1991b; Ben-
However, wet dog shakes, diarrhea, grooming and other
Eliyahu et al. 1992; Trujillo and Akil 1994; Elliott et al.
reported attenuated opiate withdrawal behaviors are
1995; Inturrisi 1997; Trujillo 1995, 1999). It is now gen-
characteristic only for adult animals and are not present
erally agreed that chronic coadministration of MK-801
in the infant rat (Jones and Barr 1995; Windh et al. 1995;
and other NMDA antagonists prevents the development
Thornton and Smith 1997; Thornton et al. 1997). The ef-
of opiate tolerance and dependence in rodents (Herman
fect of these drugs on opiate withdrawal behaviors
et al. 1995; Trujillo 1995, 1999). More controversial are
unique to infants was unknown. We found that, when be-
the reports about the effects of MK-801 and other
havioral measures appropriate for the infant rat are
NMDA antagonists on the expression of morphine with-
adopted, acute administration of NOS inhibitors effec-
drawal (Herman et al. 1995). Some laboratories report
tively reduced a variety of the opioid abstinence symp-
that NMDA antagonists inhibit withdrawal symptoms
toms in 7-day-old rats, including head moves, moving
(Brent and Chahl 1993; Cappendijk et al. 1993; Tanga-
paws, rolling together, walking, and increased quiet be-
nelli et al. 1991); others report that they do not attenuate
opiate withdrawal (Trujillo and Akil 1991a, 1991b), and
Vaupel et al. reported that, although NOS inhibitors
still others report mixed results (Rasmussen et al. 1991;
produced significant decreases in weight loss, diarrhea,
Thorat et al. 1994). At first glance, it seems that our re-
wet dog shakes and grooming in adult rats, some with-
sults are in line with the last group, because in our exper-
drawal signs (e.g., exploratory activity) were increased.
iments both MK-801 and AP-5 failed to attenuate some
They attributed this to the relatively low level of opiate
opiate withdrawal behavior in infant rats, and both inten-
dependence of the subjects due to short morphine treat-
sified several others. However, it is not clear if our nega-
tive results for MK-801 are replicating those negative re-
cally but also phylogenetically. As Herman et al. (1995)
sults from the adult or are revealing true differences be-
first pointed out, depending on species of rodent, the ef-
fect of MK-801 on opiate withdrawal is different. In
The reason for the difference between chronic treat-
guinea pigs, MK-801 is consistently reported to inhibit
ment and acute treatment of MK-801 in adult animals is
various morphine withdrawal symptoms (Tanganelli et
unclear (Trujillo 1995). There may exist fundamental
al. 1991; Brent and Chahl 1993). In adult rats, the results
differences between the mechanisms of the development
are less consistent (Rasmussen et al. 1991; Bhargava et
or acquisition of opiate dependence and the physical ex-
al. 1995). Most controversial are the effects of MK-801
pression of dependence withdrawal. Although the
in inhibiting morphine withdrawal symptoms in mice.
NMDA receptor plays a vital role in the development of
There have been reports of both significant effects
opiate dependence, once the dependence is established,
(Tanganelli et al. 1991) and observations that failed to
its expression may not rely on the functioning of the
obtain significant effects (Thorat et al. 1994). These dif-
NMDA receptor. Studies by Rasmussen and colleagues
ferences among species may indicate a relationship be-
lend support to the concept that NMDA receptors play
tween the level of brain evolution and the degree of MK-
only a minor role in the expression of opiate withdrawal.
801 response (Herman et al. 1995). In our experiments, a
In opiate-dependent rats, there is a marked increase in
possible reason for the ineffectiveness of MK-801 in in-
firing of locus coeruleus (LC) neurons during naltrex-
hibiting opiate withdrawal behaviors in the infant rat is
one-precipitated withdrawal; and opiate withdrawal in-
that neural systems of the neonatal rat are ontogenetical-
creases glutamate and aspartate efflux in the LC (Ras-
ly less evolved than that of the adult rat and phylogeneti-
mussen and Aghajanian 1989; Rasmussen et al. 1996).
cally less evolved than that of the guinea pig. Therefore,
This increase in activity of LC neurons has been hypoth-
the mechanism by which MK-801 takes effect in reduc-
esized to be important in opiate withdrawal symptoms
ing various opiate withdrawal symptoms in guinea pigs
(Rasmussen et al. 1996). The excitatory effect of gluta-
and adult rats is ineffective in neonatal rats. Whether the
mate in the LC is mediated largely through AMPA (α-
different development status of NMDA receptors is re-
amino-3-hydroxy-5-methyl-4-isoxazole propionic acid)
sponsible for these observed differences of MK-801 in in-
receptors, whereas NMDA receptors mediate little, if
hibiting opiate withdrawal in different species and differ-
any, of the withdrawal-induced excitation of the LC
ently aged rodents is unknown and deserves further study.
(Rasmussen 1995; Rasmussen et al. 1991, 1996). Re-
It is important to note that our data do not support, at
cently, this group also reported that the selective metabo-
least in the infant rat, the notion that the production of
tropic glutamate receptor (mGlu2/3 receptor) agonist
NO is linked to the activation of NMDA receptors (Mao
LY354740 attenuated morphine withdrawal induced acti-
et al. 1995; Mayer and Mao 1999) during opiate with-
vation of LC neurons and behavioral signs of morphine
drawal. When NOS inhibitors were applied, withdrawal
withdrawal (Vandergriff and Rasmussen 1999). These
behaviors were inhibited, but withdrawal syndromes
data suggest that, although NMDA receptors are impor-
were still manifest after the blockade of NMDA recep-
tant in the acquisition of morphine dependence, once the
tors by NMDA receptor antagonists. Our data thus sug-
dependence is established, NMDA receptors may play a
gest that the activation of NMDA receptors is not neces-
minor role. Other glutamate receptors such as AMPA
sarily the single trigger of the activation and production
and metabotropic glutamate receptors may play the ma-
of NO. It seems that once opiate dependence is estab-
lished, the activation of NO during withdrawal is trig-
On the other hand, the reason that we obtained nega-
gered by some factors that are “downstream” to the
tive results for the NMDA receptor antagonists in the
neonatal rat may be because NMDA receptors functiondifferently in neonatal rats than in adult rats. During de-velopment, NMDA receptors undergo qualitative and
quantitative changes (Kalb et al. 1992; Hori and Kanda1994; Kalb and Fox 1997). These include alterations in
Adams ML, Kalicki JM, Meyer ER, Cicero TJ (1993) Inhibition
density of the receptor (Morin et al. 1989; Represa et al.
of the morphine withdrawal syndrome by a nitric oxide syn-
1989; Tremblay et al. 1988) and sensitivity to magne-
thase inhibitor, NG-nitro-L-arginine methyl ester. Life Sci52:PL245–PL249
sium (Ben-Ari et al. 1988; Bowe and Nadler 1990;
Akaoka H, Aston-Jones G (1991) Opiate-withdrawal-induced hy-
Morrisett et al. 1990) of NMDA receptors during the
peractivity of locus coeruleus neurons is substantially mediat-
course of development. At the same time, the various
ed by augmented excitatory amino acid input. J Neurosci
subunits (R1, R2A–D, etc.) of the NMDA receptor com-
Barr GA (1992) Behavioral effects of opiates during development.
plex also undergo dramatic differential maturation dur-
In: Miller MW (ed) Development of the central nervous
ing the first few postnatal weeks of the animal (Pollard
system: effects of alcohol and opiates. Wiley-Liss, New York,
et al. 1993; Laurie and Seeburg 1994; Zhong et al. 1994,
1995; Guilarte and McGlothan 1998), resulting in con-
Barr GA (1993) Reinforcing properties of opiates during early de-
siderably different function of the NMDA receptor
velopment. In: Hammer RP (ed) The neurobiology of opiates. CRC Press, Boca Raton, pp 63–83
(Colwell et al. 1998). The effectiveness of MK-801 in
Barr GA, Jones KL (1994) Opiate withdrawal in the infant. Neuro-
inhibiting opiate withdrawal varies not only ontogeneti-
Barr GA, Zmitrovich A, Hamowy AS, Yan P, Liu R, Wang S,
Kimes AS, Vaupel DB, London ED (1993) Attenuation of some
Hutchings DE (1998) Neonatal withdrawal following pre- and
signs of opioid withdrawal by inhibitors of nitric oxide syn-
postnatal exposure to methadone in the rat. Pharmacol Bio-
Laurie DJ, Seeburg PH (1994) Regional and developmental het-
Bell JA, Beglan CL (1995) MK-801 blocks the expression but not
erogeneity in splicing of the rat brain NMDAR1 mRNA. J
the development of tolerance to morphine in the isolated spi-
nal cord of the neonatal rat. Eur J Pharmacol 294:289–296
Mao J, Price DD, Mayer DJ (1995) Mechanisms of hyperalgesia
Ben-Ari Y, Cherubini E, Krnjevic K (1988) Changes in voltage
and morphine tolerance: a current view of their possible inter-
dependence of NMDA currents during development. Neurosci
Marek P, Ben-Eliyahu S, Gold M, Liebskind JC (1991a) Excitato-
Ben-Eliyahu S, Marek P, Vaccarino AL, Mogil JS, Sternberg WF,
ry amino acid antagonists (kynurenic acid and MK-801) atten-
Lifebskind JC (1992) The NMDA receptor antagonist MK-
uate the development of morphine tolerance in the rat. Brain
801 prevents long-lasting no-associative morphine tolerance in
Marek P, Ben-Eliyahu S, Vaccarino AL, Liebskind JC (1991b) De-
Bhargava HN, Reddy PL, Gudehithlu KP (1995) Down-regulation
layed application of MK-801 attenuates morphine tolerance in
of N-methyl-D-aspartate (NMDA) receptors of brain regions
and spinal cord of rats treated chronically with morphine. Gen
Mayer DJ, Mao J (1999) Mechanisms of opioid tolerance: a cur-
rent view of cellular mechanisms. Pain Forum 8:14–18
Bowe MA, Nadler JV (1990) Developmental increase in the sensi-
Moore PK, Babbedge RC, Wallace P, Gaffen ZA, Hart SL (1993)
tivity to magnesium of NMDA receptors on CA1 hippocampal
7-Nitro indazole, an inhibitor of nitric oxide synthase, exhibits
pyramidal cells. Brain Res Dev Brain Res 56:55–61
anti-nociceptive activity in the mouse without increasing
Brent PJ, Chahl LA (1993) Enhancement of the opiate withdrawal
blood pressure. Br J Pharmacol 108:296–297
response by antipsychotic drugs in guinea pigs is not mediated
Morin AM, Hattori H, Wasterliain CG, Thomson D (1989)
by sigma binding sites. Eur Neuropsychopharmacol 3:23–32
[H3]MK-801 binding sites in neonate rat brain. Brain Res
Cappendijk SL, Vries R, Dzoljic MR (1993) Inhibitory effect of
nitric oxide (NO) synthase inhibitors on naloxone-precipitated
Morrisett RA, Mott DD, Lewis DV, Wilson WA, Swartzwelder HS
withdrawal syndrome in morphine-dependent mice. Neurosci
(1990) Reduced sensitivity of the N-methyl-D-aspartate com-
ponent of synaptic transmission to magnesium in hippocampal
Carden SE, Barr GA, Hofer MA (1991) Differential effects of spe-
slices from immature rats. Brain Res Dev Brain Res
cific opioid receptor agonists on rat pup isolation calls. Brain
Naeye KL, Blanc W, Leblank W, Khatamee MA (1973) Fetal
Colwell CS, Cepeda C, Crawford C, Levine MS (1998) Postnatal
complications of maternal heroin addiction: abnormal growth,
development of glutamate receptor-mediated responses in the
infections, and episodes of distress. J Pediatr 83:1055–1061
Pollard H, Khrestchatisky M, Moreau J, Ben Ari Y (1993) Tran-
Connaughton JF, Reeser D, Schut J, Finegan LP (1977) Perinatal
sient expression of the NR2C subunit of the NMDA receptor
addiction: outcome and management. Am J Obstet Gynecol
in developing rat brain. Neuroreport 4:411–414
Rasmussen K (1995) The role of the locus coeruleus and NMDA
Elliott K, Kest B, Man A, Kao B, Inturrisi CE (1995) N-Methyl-D-
and AMPA receptors in opiate withdrawal. Neuropsychophar-
aspartate (NMDA) receptors, mu and kappa opioid tolerance,
and perspectives on new analgesic drug development. Neuro-
Rasmussen K, Aghajanian GK (1989) Withdrawal-induced activa-
tion of locus coeruleus neurons in opiate-dependent rats: atten-
Fitzgerald M (1995) Developmental biology of inflammatory pain.
uation by lesions of the nucleus paragigantocellularis. Brain
Geller LM, Geller ES (1966) A simple technique for permanent
Rasmussen K, Fuller RW, Stockton ME, Perry KW, Swinford RM,
marking of newborn albino rats. Psychol Rep 18:221–222
Ornstein PL (1991) NMDA receptor antagonists suppress be-
Guilarte TR, McGlothan JL (1998) Hippocampal NMDA receptor
haviors but not norepinephrine turnover or locus coeruleus
mRNA undergoes subunit specific changes during develop-
unit activity induced by opiate withdrawal. Eur J Pharmacol
mental lead exposure. Brain Res 790:98–107
Herman BH, Vocci F, Bridge P (1995) The effects of NMDA re-
Rasmussen K, Kendrick WT, Kogan JH, Aghajanian GK (1996) A
ceptor antagonists and nitric oxide synthase inhibitors on opio-
selective AMPA antagonist, LY293558, suppresses morphine
id tolerance and withdrawal. Neuropsychopharmacology
withdrawal-induced activation of locus coeruleus neurons and
behavioral signs of morphine withdrawal. Neuropsychophar-
Hori Y, Kanda K (1994) Developmental alterations in NMDA re-
ceptor-mediated [Ca2+] elevation in substantia gelatinosa neu-
Represa A, Tremblay E, Ben-Ari Y (1989) Transient increase of
rons of neonatal rat spinal cord. Brain Res Dev Brain Res
NMDA-binding sites in human hippocampus during develop-
Hutchings DE (1987) Drug abuse during pregnancy: embryopathic
Scalzo FM, Burge LJ (1994) The role of NMDA and sigma sys-
and neurobehavioral effects. In: Braude MC, Zimmermam AM
tems in the behavioral effects of phencyclidine in preweanling
(eds) Genetic and perinatal effects of abused substances. Aca-
Tanganelli S, Antonelli T, Morari M, Bianchi C, Beani L (1991)
Inturrisi CE (1997) Preclinical evidence for a role of glutamatergic
Glutamate antagonists prevent morphine withdrawal in mice
systems in opioid tolerance and dependence. Semin Neurosci
and guinea pigs. Neurosci Lett 122:270–272
Thorat SN, Barjavel M, Matwyshyn GA, Bhargava HN (1994)
Jones KL, Barr GA (1995) Ontogeny of morphine withdrawal in
Comparative effects of NG-monomethyl-L-arginine and MK-
801 on the abstinence syndrome in morphine-dependent mice.
Kalb RG, Fox AL (1997) Synchronized overproduction of AMPA,
kainate, and NMDA glutamate receptors during human spinal
Thornton SR, Smith FL (1997) Characterization of neonatal rat
cord development. J Comp Neurol 384:200–210
fentanyl tolerance and dependence. J Pharmacol Exp Ther
Kalb RG, Lidow MS, Halsted MJ, Hockfield S (1992) N-Methyl-
D-aspartate receptors are transiently expressed in the develop-
Thornton SR, Wang AF, Smith FL (1997) Characterization of neo-
ing spinal cord ventral horn. Proc Natl Acad Sci USA
natal rat morphine tolerance and dependence. Eur J Pharmacol
Tremblay E, Roisin MP, Represa A, Charriaut-Marlangue C, Ben-
Vaupel DB, Kimes AS, London ED (1995a) Comparison of 7-ni-
Ari Y (1988) Transient increased density of NMDA binding
troindazole with other nitric oxide synthase inhibitors as atten-
sites in the developing rat hippocampus. Brain Res 461:393–
uators of opioid withdrawal. Psychopharmacology 118:361–
Trujillo KA (1995) Effects of noncompetitive N-methyl-D-aspar-
Vaupel DB, Kimes AS, London ED (1995b) Nitric oxide synthase
tate receptor antagonists on opiate tolerance and physical de-
inhibitors: preclinical studies of potential use for treatment of
pendence. Neuropsychopharmacology 13:301–307
opiate addiction. Neuropsychopharmacology 13:315–322
Trujillo KA (1999) Cellular and molecular mechanisms of opioid
Wilson GS, McCreary R, Kean J, Baxter JC (1979) The develop-
tolerance and dependence: progress and pitfalls. Pain Forum
ment of preschool children of heroin-addicted mothers: a con-
Trujillo KA, Akil H (1991a) Inhibition of morphine tolerance and
Windh RT, Little PJ, Kuhn CM (1995) The ontogeny of mu opiate
the dependence by the NMDA receptor antagonist MK-801.
tolerance and dependence in the rat: antinociceptive and bio-
chemical studies. J Pharmacol Exp Ther 273:1361–1374
Trujillo KA, Akil H (1991b) Opiate tolerance and dependence:
Zhong J, Russell SL, Pritchett DB, Molinoff PB, Williams K
recent findings and synthesis. New Biol 3:915–923
(1994) Expression of mRNAs encoding subunits of the N-
Trujillo KA, Akil H (1994) Inhibition of opiate tolerance by non-
methyl-D-aspartate receptor in cultured cortical neurons. Mol
competitive N-methyl-D-aspartate receptor antagonists. Brain
Zhong J, Carrozza DP, Williams K, Pritchett DB, Molinoff PB
Vandergriff J, Rasmussen K (1999) The selective mGlu2/3 recep-
(1995) Expression of mRNAs encoding subunits of the
tor agonist LY354740 attenuates morphine-withdrawal-
NMDA receptor in developing rat brain. J Neurochem
induced activation of locus coeruleus neurons and behavioral
signs of morphine withdrawal. Neuropharmacology 38:217–222
Myeloma group MELPHALAN, PREDNISOLONE AND THALIDOMIDE (MPT) Summary of changes (2013): Tabulation of dose modification Change of emetic risk INDICATION As initial therapy or at relapse in patients thought unsuitable for CTD / autografting. PRE-ADMINISTRATION 1. Ensure all the following staging investigations are done: o FBC & film o Clotting screen o U&Es
C H A P T E R • SIGNAL TRANSDUCTION PATHWAYS Signal Transduction Pathways Organization Receptors Soluble Receptors Transmembrane Receptors Enzyme Coupled Receptors G-Protein Coupled Receptors Ion-Channel Coupled Receptors Second Messengers Amplifiers Integrators Inhibitors • • • • • • • • • • • • SIGNAL TRANSDUCTION PATHWAYS
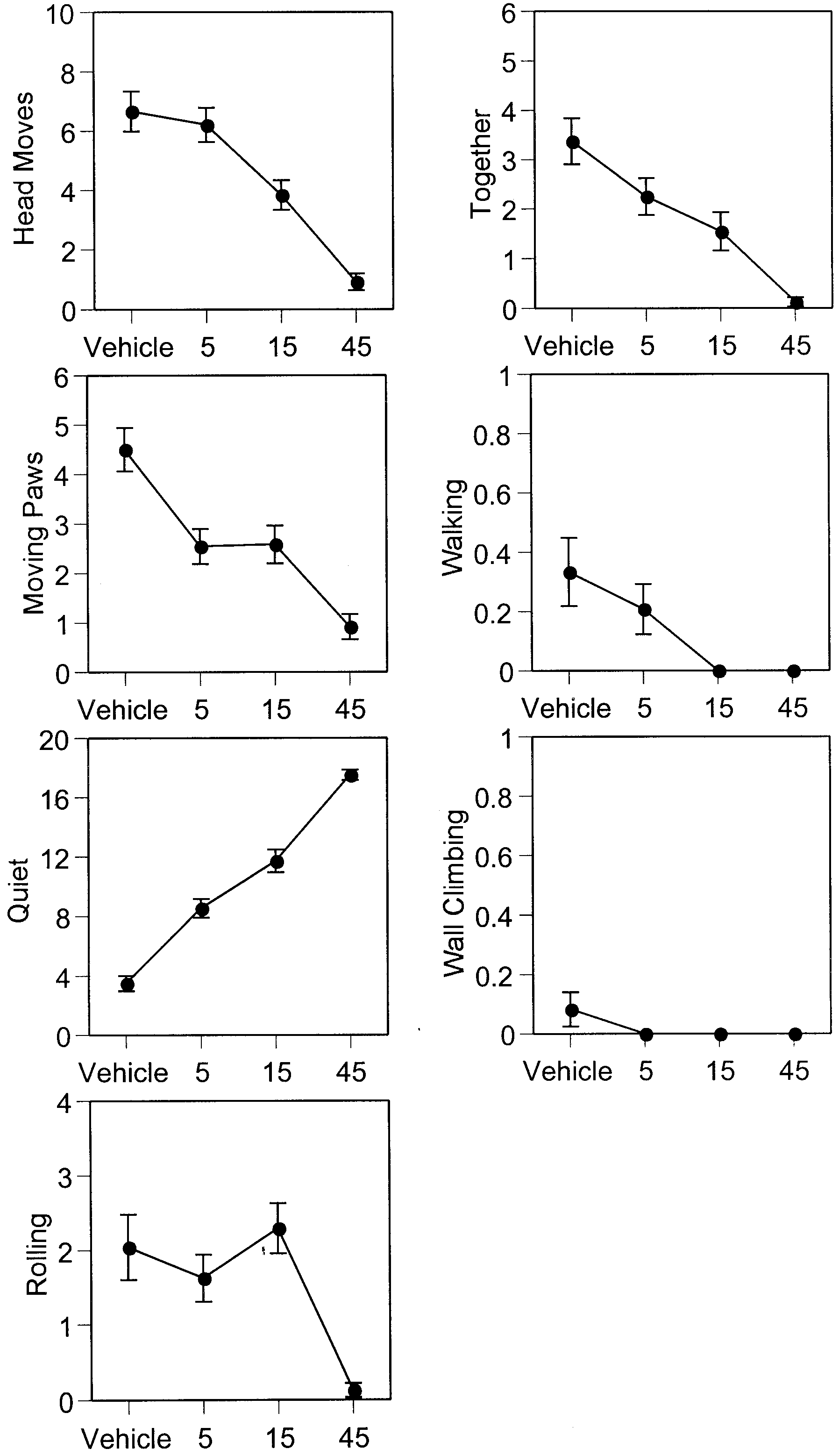
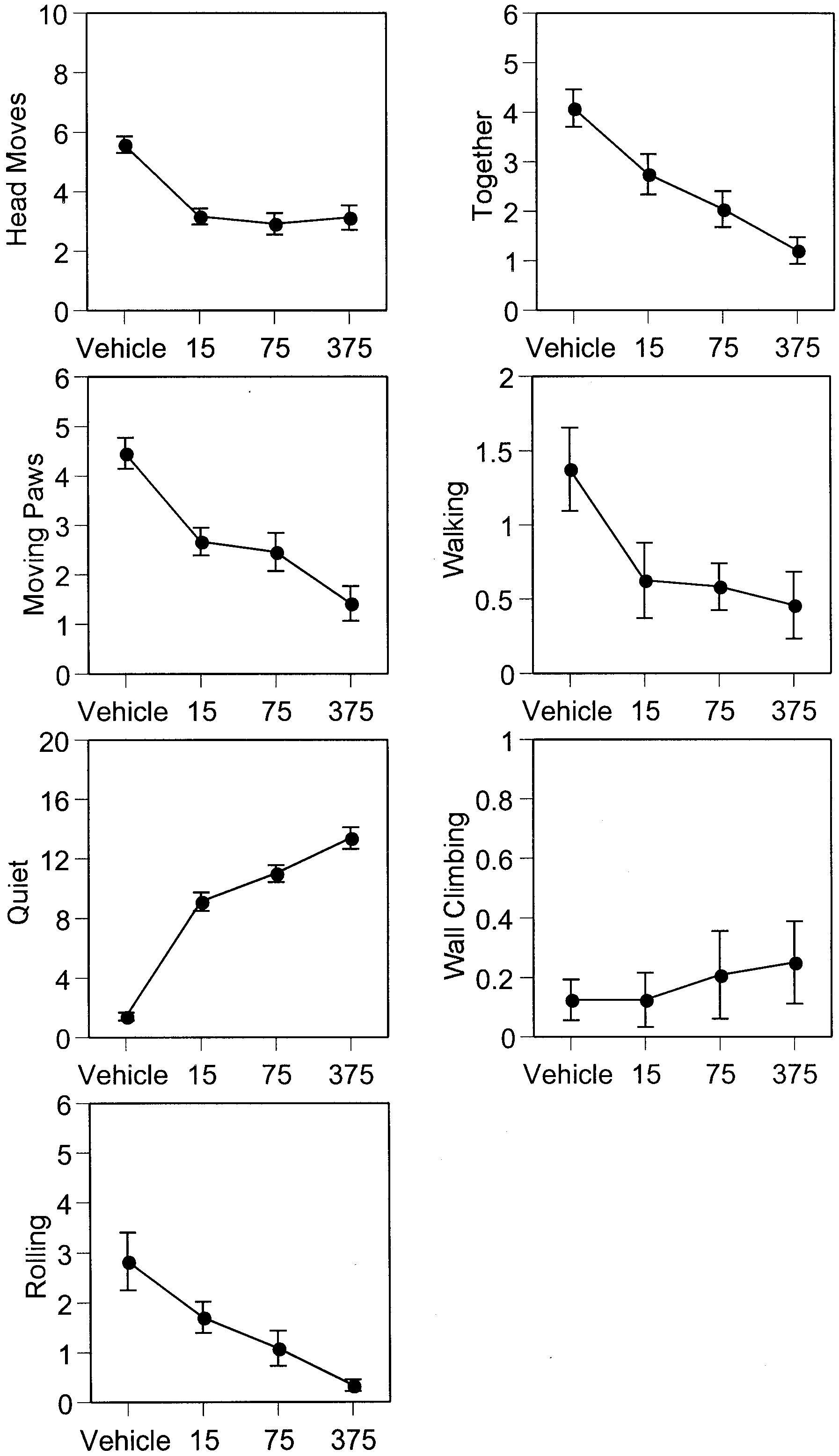 Fig. 1 Dose effect of 7-NI. Ordinate mean occurrences (mean ±
Fig. 1 Dose effect of 7-NI. Ordinate mean occurrences (mean ±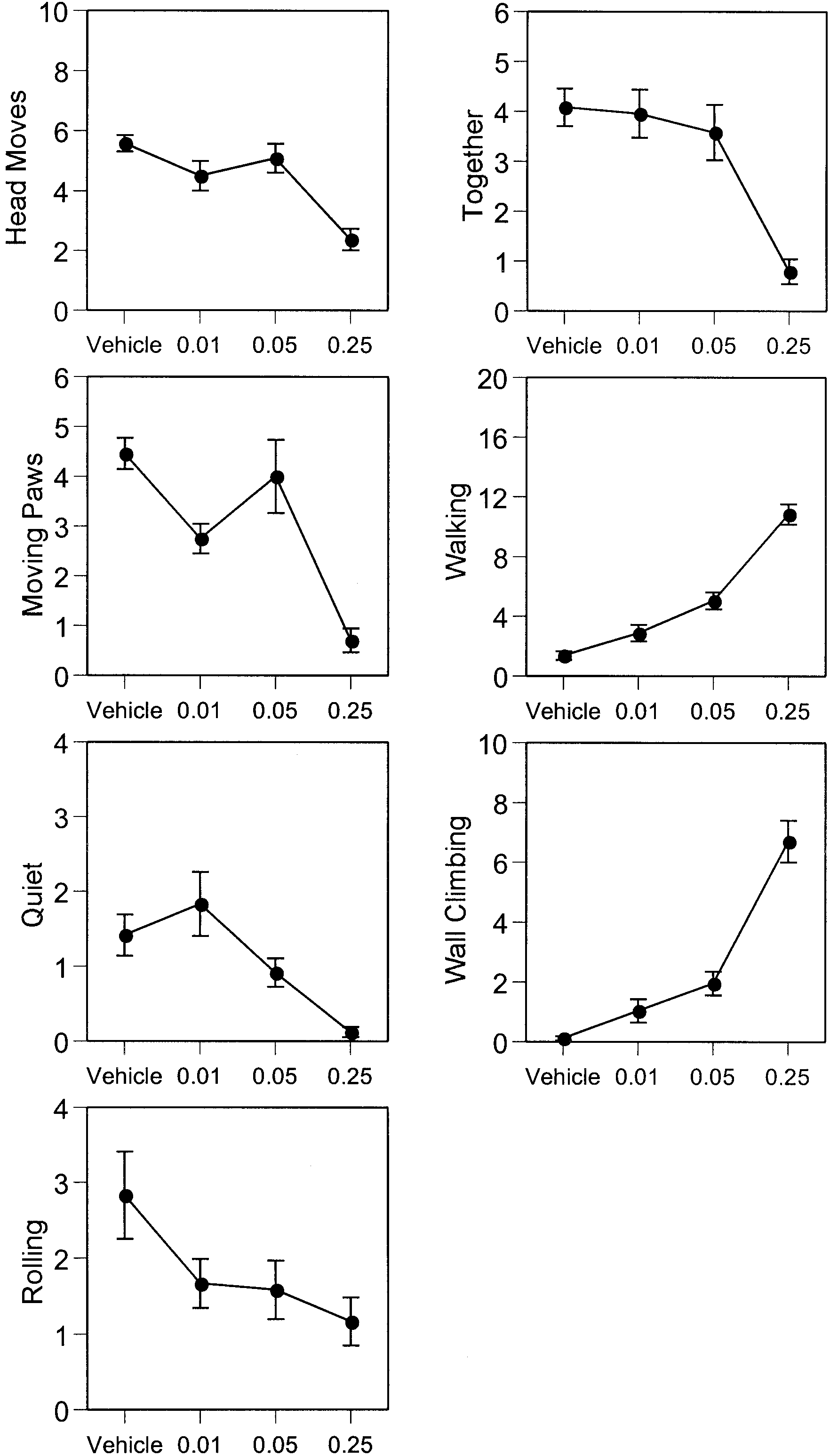
 Fig. 3 Dose effect of MK-801. Ordinate mean occurrences (mean
Fig. 3 Dose effect of MK-801. Ordinate mean occurrences (mean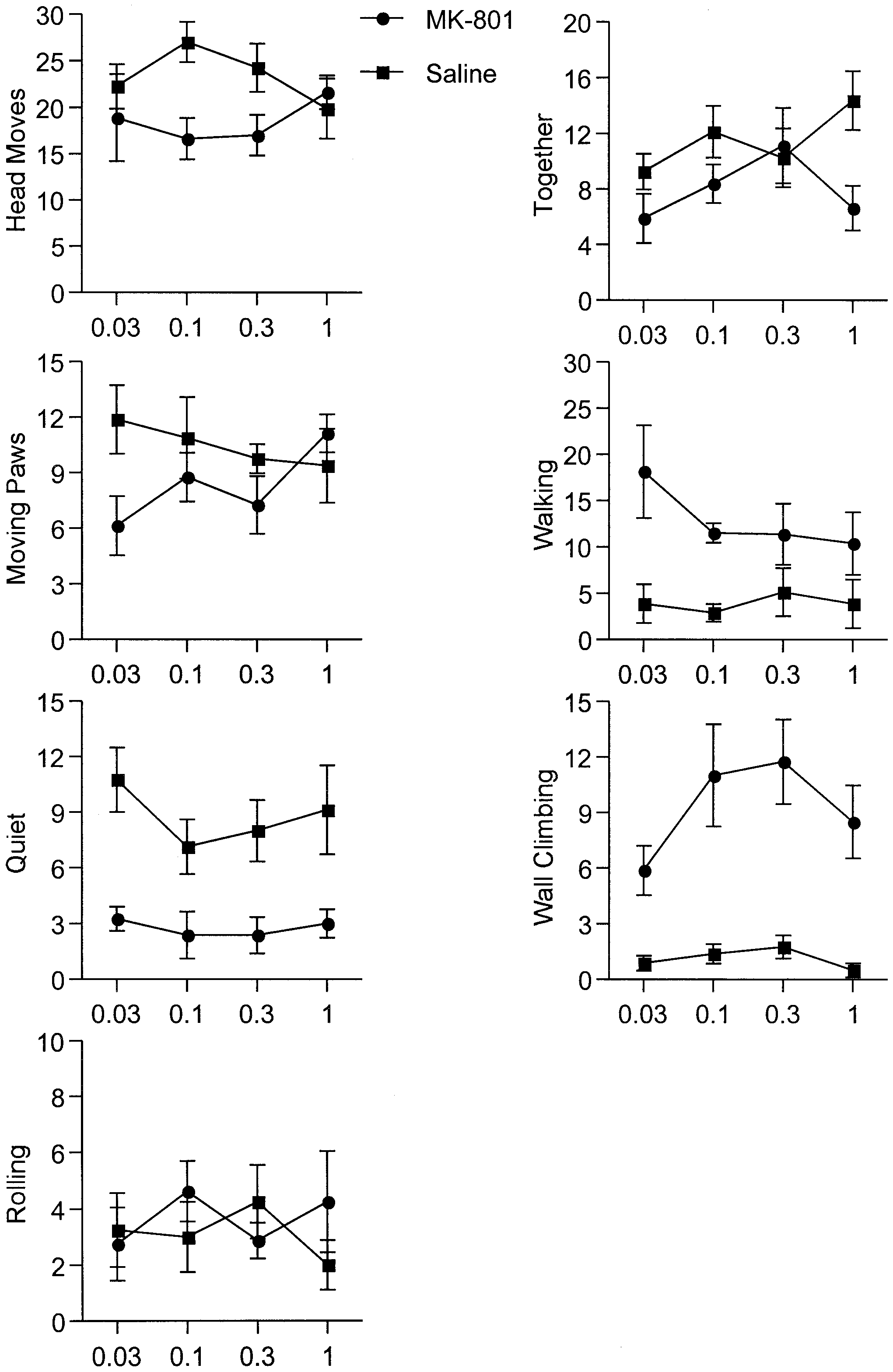 Table 6 Results of the analysis of the interaction between MK-
Table 6 Results of the analysis of the interaction between MK-