Veterinary Parasitology 101 (2001) 405–414
Molecular approaches to studying benzimidazole
J.F. Humbert a,∗, J. Cabaret b, L. Elard c, V. Leignel d, A. Silvestre b
a INRA, Station d’Hydrobiologie Lacustre, BP 511, 74203 Thonon Cedex, France
b INRA, Station de Pathologie Aviaire et de Parasitologie, 37380 Nouzilly, France
c Laboratoire de Parasitologie Fondamentale et Fonctionnelle,Université de Paris VI, 75252 Paris Cedex 05, France
d Laborato´ıre de Biolog´ıe et de Génétique Evolutive, Faculté des Sciences et Techniques,Abstract
Molecular techniques are of growing importance in the study of anthelmintic resistance in tri-
chostrongylid worm populations. A knowledge of the genetic determinants of benzimidazole (BZ)resistance has made it possible to construct a molecular tool for genotyping individual worms,in respect of mutation of the -tubulin gene responsible for BZ resistance. This tool offers newpossibilities in the diagnosis of BZ resistance, and also in the study of anthelmintic use and otherbreeding management factors that can affect the selection of BZ-resistant alleles in worm popu-lations. New molecular methods have also made it possible to study the origin and diversity ofBZ-resistant alleles in trichostrongylid populations. The results demonstrate the value of a multi-disciplinary approach to the study of anthelmintic resistance, combining molecular, ecological andepidemiological techniques. 2001 Elsevier Science B.V. All rights reserved. Keywords: Nematodes; Trichostrongilidae; Benzimidazole; Resistance; Molecular techniques
1. Introduction
As with bacteria and antibiotics, or with insects and insecticides, the trichostrongylid ne-
matode parasites of domestic small ruminants (sheep and goats) have acquired resistance toanthelmintics. Although, this development affects all available anthelmintics, the benzimi-dazoles (BZs) present the most pressing problem. These drugs have been widely used sincethe 1960s (see review on goats in Cabaret, 2000), mostly in lactating ruminants, because of
∗ Corresponding author. Tel.: +33-450-26-7809; fax: +33-450-26-0760. E-mail address: humbert@thonon.inra.fr (J.F. Humbert).
0304-4017/01/$ – see front matter 2001 Elsevier Science B.V. All rights reserved. PII: S 0 3 0 4 - 4 0 1 7 ( 0 1 ) 0 0 5 6 5 - 9
J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
legal constraints on the use of anthelmintics based on residues in milk. A recent investiga-tion showed that BZs represented more than 80% of all treatments against nematodes usedin French dairy goats (Hoste et al., 2000). This has prompted many attempts to identify thegene responsible for the BZ resistance.
These studies have demonstrated that BZ resistance in trichostrongylid nematodes is
principally linked to alterations in the gene that encodes for -tubulin, the target site ofthe BZs (Roos et al., 1990; Geary et al., 1992; Kwa et al., 1993a; Beech et al., 1994;Lubega et al., 1994; Grant and Mascord, 1996). A mutation causing BZ resistance inthe three main gastrointestinal species (Teladorsagia circumcincta, Trichostrongylus col-ubriformis and Haemonchus contortus) was identified at amino acid 200 of the isotype1 -tubulin gene (Kwa et al., 1993b, 1994, 1995; Elard et al., 1996; Elard and Humbert,1999).
This knowledge of the molecular basis of the resistance to xenobiotics has allowed the de-
velopment of PCR-based methods for their detection. With the expanding use of moleculartechniques for diagnosis, these methods are becoming increasingly important in bacteriol-ogy (Maggs et al., 1998). Our team has developed a PCR-based tool for the diagnosis of BZresistance in worm populations (Humbert and Elard, 1997; Elard et al., 1999). In additionto its diagnostic value, this tool makes it possible to study resistance traits more generally. The putative cost of the mutation has, for example, implications for the reversibility ofresistance, and for breeding management practices in relation to the selection of resistantworms.
This paper compares the efficacy of this molecular tool with classical methods for diag-
nosing BZ resistance, and shows how molecular studies can be a useful means of under-standing the acquisition and development of resistance to BZ in a parasite population (asingle species of nematodes) or in a community (several species of nematodes). 2. A molecular tool for the diagnosis of BZ resistance
The mutation involved in BZ resistance causes the replacement of a phenylalanine by
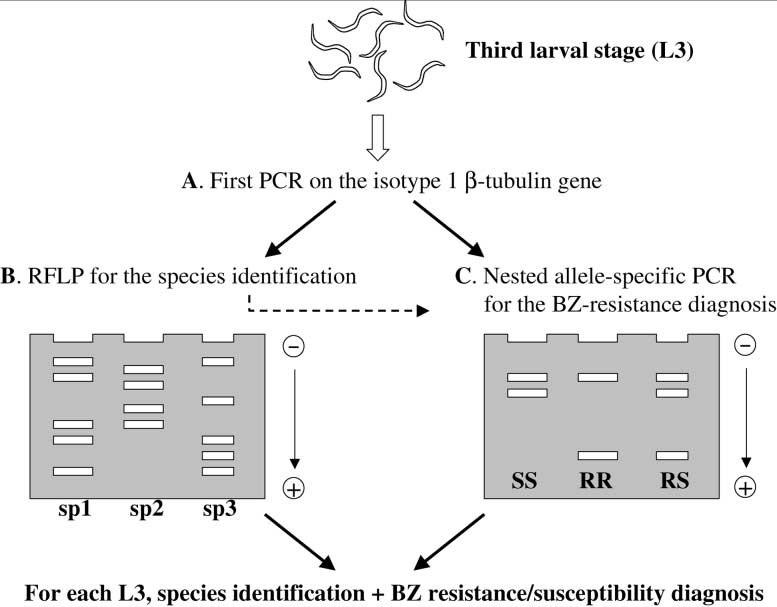
a tyrosine at residue 200 of the isotype 1 -tubulin gene. We have developed, for the firsttime, an allele-specific PCR which makes it possible to determine the genotype (Phe/Phe,Phe/Tyr and Tyr/Tyr) of adult T. circumcincta worms in relation to the nature of residue200 (Fig. 1) (Humbert and Elard, 1997; Elard et al., 1999). This method has also beenmodified for use on the third larval stage (L3) to allow the identification of the threemost prevalent trichostrongylid species (T. circumcincta, T. colubriformis and H. contor-tus) and to diagnose their BZ resistance or susceptibility (Fig. 2) (Silvestre and Humbert,2000).
This molecular tool can be used to estimate the proportion of each genotype (rr: resistant;
rS and SS: both susceptible) in a trichostrongylid worm community. The level of BZ resis-tance in a worm population can be defined as the proportion of homozygous mutant worms(Tyr/Tyr = rr), because these are the only ones which survive a recommended dose of BZ(Elard and Humbert, 1999). Thus, BZ resistance can be described quantitatively rangingfrom 0 to 100%. The advantages of this method over classical approaches will be discussedin the next section. J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
Fig. 1. Principle of allele-specific PCR for the genotyping of worms in respect of residue 200 of their isotype 1-tubulin gene: (A) detection of BZ-susceptible allele; (B) detection of BZ-resistant allele. This allele-specificPCR contains four primers: a and b are BZ-non-specific primers; c is a BZ-susceptible primer; d is a BZ-resistantprimer. Three fragments can thus be amplified: a BZ-non-specific fragment (1); a BZ-susceptible fragment (2); and aBZ-resistant fragment (3). In homozygous TTC/TTC worms only two fragments (1 and 2) are detected in agaroseelectrophoresis. In homozygous TAC/TAC worms, only two fragments (2 and 3) are detected in agarose elec-trophoresis. In heterozygous TTC/TAC worms, the three fragments (1–3) are detected in agarose electrophoresis.
Fig. 2. Principle of the technique used for species identification and BZ resistance diagnosis in the third larval stageof the three principal trichostrongylid species: T. circumcincta; T. colubriformis; and H. contortus. (A) Individualthird stage larvae were crushed and a first PCR used to amplify a large fragment of the isotype 1 -tubulin gene. (B) Species identification was performed by enzymatic digestion. (C) Nested allele-specific PCR was performedwith a set of primers for each species. This reveals the BZ resistance or -susceptibility of each larva. J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–4143. Comparison of our molecular tool with classical methods used to estimate BZ resistance
We compared our molecular tool for diagnosing BZ resistance with three other widely
used methods: the fecal egg count reduction test (FECRT), which estimates the fall in eggexcretion after a BZ treatment; the egg hatch assay (EHA), which makes it possible toestimate a lethal dose 50 (the killing of 50% of eggs); and the adult reduction test (ART)which estimates the reduction in the adult worm population after treatment (for details ofthese tests, see Coles et al., 1992; Wood et al., 1995). The first two tests give an estimateof the resistance of the whole worm community, while the ART determines the resistancewithin each species. The latter method requires a necropsy and has mostly been used inexperimental designs rather than in resistance surveys. The first advantage of our approachis that it allows an estimation of BZ resistance for the three most prevalent trichostrongylidspecies (T. circumcincta, T. colubriformis and H. contortus) using infective larvae, whichcan be readily obtained from fecal cultures. The second advantage of this molecular methodis that it detects the onset of resistance, e.g., when resistant individuals in a worm populationfirst appear. The number of worms that have to be processed to find at least one resistantindividual is 100, 50, 35, and 20 for, respectively, 4, 8, 10 and 12% resistant genotypesin the population (Elard et al., 1999). Conversely, FECRT and EHA detected resistanceunequivocally only when 50% or more worms (T. colubriformis and T. circumcincta) wereresistant; neither method detected resistance below 25% (Martin et al., 1989).
In the estimation of resistance to BZ, there was generally a significant correlation be-
tween our molecular typing and two classical approaches (EHA and ART), but not with theFECRT (Table 1). The coefficient of determination (the part of the variance explained bythe relationship between the results obtained by two techniques) remained modest and wasat best lower than 50% (data not shown). The FECRT (from the data we reanalyzed) wasnot consistently correlated with EHA and ART, possibly because of differences in the wormcommunities and in the technique used to calculate it (either derived from Coles et al., 1992or Wood et al., 1995). As we summarized here, the FECRT calculations are very differ-ent (using four or two batches of hosts and estimating fecal egg counts from geometric orarithmetic means). Hosts are evaluated on fecal egg counts at the moment of treatment (T1)and/or only 10–14 days afterwards (T2). Untreated controls are checked at the same times(C1 and C2, respectively). We have then the following possibilities in calculating FECRT:
Coles et al. (1992): 100 × (1 − T2/C2). Dash et al. (1988): 100 × (T2/T1) × (C1/C2). Presidente (1985): 100 × (1 − (log T2/log T1)(log C1/log C2)). Before/after evaluation, the hosts serving as their own control: 100 × (T2 − T1)/T1.
The calculated FECRT yields very different estimates of efficacy and hence, of resis-
tance, as shown by, among others, Kochapakdee et al. (1995) when using treatment withfenbendazole: 18, 30, 59, and 25%, respectively. This might in part explain the inconsistencybetween FECRT and the other methods of estimating resistance. FECRT is highly integra-tive of: community structure (in terms of species, Cabaret et al., 1995); intensity of infection(Marriner and Bogan, 1981: efficacy of BZs is highly reduced in heavily infected hosts);timing of dosing (Hennessy et al., 1993: efficacy is increased in fasting lambs); and breeding
J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
Table 1Comparison of techniques used to detect BZ resistance (original data reanalyzed)
a Fecal egg count reduction test. b Egg hatch assay. c Adult reduction test. d Molecular typing.
management (Silvestre et al., 2000). The FECRT information covers resistant genotypes aswell as susceptible genotypes (of all species) which escaped from the drug. The informationobtained at a given site or farm might be highly misleading if performed on a single occasionand without further in vitro tests such as the EHA or our genotyping procedure. 4. Molecular techniques for studying the origin of the BZ resistance alleles in worm populations
One critical question about BZ resistance concerns the origin of the BZ resistance alleles
in worm populations. These alleles may arise by spontaneous mutation or by migrationbut they may also have been present for a long time as a rare allele. Studies with othermodels of resistance to xenobiotics demonstrated that migration plays a fundamental rolein such things as, the dispersion of insecticide resistance genes in mosquitoes (Raymond etal., 1991), and of antibiotic resistance among some species of bacteria (O’Brien, 1997). Insuch cases, measures can be adopted to limit the spread of the resistance alleles. But, whenthe resistance allele is present as a rare allele in a population, the spread of resistance ismore difficult to prevent because the use of that drug will inevitably constitute a selectionpressure in favor of this allele.
There have been few studies on the origin of the BZ resistance alleles in worm pop-
ulations. Using RFLP studies on the isotype 1 -tubulin gene, Kwa et al. (1993b) foundvarious BZ-resistant alleles in different resistant populations of H. contortus. Using the same
J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
approach on two BZ-resistant populations, Beech et al. (1994) found that the BZ resistancealleles were probably already present in two H. contortus populations before this class ofdrugs was even developed. An indirect insight into the origin of the BZ resistance allelesin parasite populations has come from Blouin et al. (1995). In a genetic population studyon the ND4 gene of the mtDNA, they showed that the worm populations of sheep in theUSA were characterized by high genetic flows between them; this can favor the spread ofresistant alleles.
By sequencing a fragment of the -tubulin gene, we have studied the diversity of BZ
resistance alleles in several goat farms located in France. Their breeding management wascharacterized by a low rate of exchange of animals between the farms and so also ofgene flow among their nematodes (Cabaret and Gasnier, 1994). In T. circumcincta andH. contortus, various resistant alleles were observed; in T. colubriformis, the same allele waspresent in all resistant populations studied (Silvestre, 2000). With regard to the demographiccharacteristics of these species, and to breeding management practices, we conclude thatsome resistant alleles have been present as rare alleles, for a long time, in all the three species,but that spontaneous mutations may also have been selected, probably in T. circumcinctaand H. contortus populations.5. Molecular tools for studying the development of BZ resistance in a worm population
The selection of a resistant allele in a worm population may occur more or less rapidly,
depending on the relative fitness of resistant and susceptible worms and also on breedingmanagement practices (see the model of Barnes et al., 1995).
Using our molecular tool, we have experimentally compared some fitness-related traits
in BZ-resistant (rr) and susceptible (rS or SS) worms of T. circumcincta. No significantdifferences were found in the fitness of these three genotypes (Elard et al., 1998). Thisobservation has recently been confirmed in the field during 2 years experiment in which novariation in the proportion of each genotype (rr, rS and SS) was found in an untreated wormpopulation (Leignel, 2000).
The consequences are important for the control of BZ resistance because relaxed selection
pressures would have been effective only if we had found evidence of a lower fitness in theresistant worms. This means that a long term use of other anthelmintic (classes) treatmentsin BZ-resistant populations does not restore susceptibility to BZ.
Breeding management practices (the use of pasture, and treatment regimen) are impor-
tant in the establishment of resistance as shown from simulations (Gettinby et al., 1989). Under-dosing has also been identified as a major factor in the establishment of resistance,although no data have been provided to back this statement. We recently tested the role of BZunder-dosing in the development of BZ resistance. With the recommended dose, only ho-mozygous resistant worms (rr) survived the BZ treatment. But, we found that in under-dosingconditions, heterozygous worms (rS) were a little less susceptible to BZ than homozygousworms (SS). Mathematical simulations have demonstrated that this small difference inBZ-susceptibility between heterozygous (rS) and homozygous worms has important con-sequences for the speed with which BZ-resistant alleles are selected (Silvestre et al., 2001). J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
We also tested the influence of anthelmintic use on the selection of BZ-resistant worms
(Leignel, 2000). Mathematical simulations suggest that alternating or combined use ofdifferent drugs can slow the development of resistance in worm populations (Barnes et al.,1995). At the end of the simulation, there was little resistance with the mixing strategy,but in the rotation strategy there was substantial resistance. Smith (1990), using simulation,also suggested that alternating anthelmintics with different modes of action may be a lesseffective resistance management strategy than administering the same drugs simultaneously. In experimentally infected paddocks (Leignel, 2000), we have found that the alternatinguse of Levamisole and BZ treatment had a limited effect on the rapidity of the selection ofBZ-resistant worms (rr) in a population initially harboring 25% of resistant worms. 6. Discussion and perspectives
Molecular techniques have made it possible to develop a tool for the species identifica-
tion of the free living stages of the parasites (from egg to infective L3 stage) and, at thesame time, to diagnose resistance to BZ. In the future, this kind of molecular tool shouldbe further developed, for example within the framework of DNA chip technology. DNAchips are solid supports on which numerous cDNA oligonucleotides can be arrayed. Theypermit notably the detection of point mutations, and of gene deletions or insertions (see DeBenedetti et al., 2000). With a better understanding of the genetic determinant of the imi-dazothiazole and avermectin drugs, it is reasonable to imagine that DNA chips will allowspecies identification and the diagnosis of drug resistance in the free living stages of allparasites of economic importance.
Molecular techniques will also provide tools for the study of the development of resis-
tance to BZ anthelmintics in worm populations. For the present, numerous questions onthose factors promoting the development of BZ resistance still need to be answered. Wehave focused mostly on the factors that give rise to increasing resistance, and have paidlittle attention to those factors that hinder its development. In this context, one of the mostintriguing questions is why in a few farms, from the same region and with a similar cli-mate and breeding management, BZ-resistant alleles remained present to a limited extent(<1%) despite the use of two or three BZ treatments per year. An integrated approachwith detailed studies of breeding management practices and of the dynamics of the threegenotypes (rr, rS and SS) might identify the key factors involved in the selection of BZresistance.
Another area in which molecular approaches to BZ resistance would be useful is the
evaluation of strategies to reduce anthelmintic use or to substitute susceptible strains forresistant ones. In situations where no effective drug remains for controlling nematodes, thelatter solution will be the only one available (Van Wyk and van Schalkwyk, 1990). Theproblem in this particular case was to assess the result of substitution, because classicaltechniques were able to detect only 25–50% of resistant worms within the community. PCR-based diagnostics would be of great utility. When the situation is not as catastrophicas the one described by Van Wyk et al. (1997) in South Africa, a critical use of the existingdrug can be planned. This involves the selective use of anthelmintics, either restricted to acategory of host (the most susceptible, such as lambs intended for meat production or goats
J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
in their first-lactation) or to selected individuals. The second solution appears very attractive,as the treatment of a few highly infected hosts should greatly reduce selective pressure on theanthelmintic. The only difficulty is in detecting highly infected hosts. Evaluation by repeatedfecal egg count is ruled out for economic reasons; in South Africa, where the blood suckingH. contortus is commonly present, the Famacha system has been used. This system isbased on a color chart with five color categories that are compared with that of the mucousmembranes of the eyes of the sheep. The animal is then scored from severely anemic tonon-anemic and those animals considered in danger of succumbing to H. contortus infectionare treated. The use of this method considerably reduces the number of treatments required(Van Wyk, personal communication, 1999) and very likely slows down the developmentof anthelmintic resistance. The method is valid in sites or at periods when H. contortus isthe main parasite. Another solution suitable for temperate areas is to detect hosts at risk,possibly by their growth rate and/or diarrhea scores (Cabaret, unpublished data). Selectivetreatment of hosts is a method that can be implemented rapidly, if the characteristics ofseverely infected hosts can be detected. Selection for host resistance to infection, on theother hand, is a long-term project. The value of such a susceptible host-based treatmentscheme could be evaluated only by means of the molecular technique we have developed;the classical techniques are very poor at detecting low level resistance (Martin et al., 1989),and PCR-based techniques can help accurately monitor any change in resistance, even atthe low levels which are the most important, when trying to control resistance. Resistancein nematodes of the digestive-tract of ruminants is probably one of the biggest challengesconfronting veterinarians and breeders. It is a challenge for production (ruminants), welfare(ruminants and horses) and zoonotic responsibility (several Trichostrongylus spp. are foundin humans from the tropics, and their elimination may depend on the same BZ drugs). Acknowledgements
The excellent technical help of C. Sauvé and J. Cortet is acknowledged. P. Dorny
(Gent Veterinary Faculty, Belgium) kindly provided unpublished data on H. contortus. A French–South Africa joint program offered the opportunity of a wider insight into theextent of the resistance problem and possible resources to fight it. Discussions with J. VanWyk (Onderstepoort Veterinary Institute, Republic of South Africa) played an importantpart in reaching this insight. The organic farming project in Auvergne (INRA-Theix) madeit possible for us to make preliminary tests of the simple detection (Famacha or diarrheascores) of nematode-susceptible lambs. Most of the data on resistance presented here wereobtained with the financial help of “Bureau des Ressources Génétiques”, “Biotechnocentre”in French Centre Region, and French Ministry of Research (Ph.D. grants to LE and VL). The English text was edited by Geoff Watts and Owen Parkes. References
Barnes, E.H., Dobson, R.J., Barger, I.A., 1995. Worm control and anthelmintic resistance adventures with a model. J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
Beech, R.N., Pritchard, R.K., Scott, M.E., 1994. Genetic variability of the -tubulin genes in
benzimidazole-susceptible and -resistant strains of Haemonchus contortus. Genetics 138, 103–110.
Blouin, M.S., Yowell, C.A., Courtney, C.H., Dame, J.B., 1995. Host movement and the genetic structure of
populations of parasitic nematodes. Genetics 141, 1007–1014.
Cabaret, J., 2000. Anthelmintic resistance in goats: from fiction to facts. In: Proceedings of the Seventh International
Cabaret, J., Gasnier, N., 1994. Farm history and breeding management influences on the intensity and specific
diversity of nematode infection of dairy goats. Vet. Parasitol. 53, 219–232.
Cabaret, J., Baudet, H.M., Devos, J., Hubert, J., Cortet, J., Sauvé, C., 1995. Studies on multispecific resistance of
gastrointestinal nematodes to benzimidazoles on dairy-goat farms. Vet. Parasitol. 60, 331–337.
Coles, G.C., Bauer, C., Borgsteede, F.H.M., Geerts, S., Klei, T.R., Taylor, M.A., Waller, P.J., 1992. World
Association for the Advancement of Veterinary Parasitology (WAAVP) methods for the detection ofanthelmintic resistance in nematodes of veterinary importance. Vet. Parasitol. 44, 35–44.
Dash, K.M., Hall, E., Barger, I.A., 1988. The role of arithmetic and geometric mean worm egg counts in faecal
egg count reduction tests and in monitoring strategic drenching programs in sheep. Aust. Vet. J. 65, 66–68.
De Benedetti, V.M., Biglia, N., Sismondi, P., De Bortoli, M., 2000. DNA chips: the future of biomarkers. Int. J.
Dorny, P., Symoens, C., Jalila, A., Vercruysse, J., Sani, R., 1995. Strongyle infections in sheep and goats under
the traditional husbandry system in peninsular Malaysia. Vet. Parasitol. 56, 121–136.
Elard, L., Humbert, J.F., 1999. Importance of the mutation of amino acid 200 of the isotype 1 -tubulin gene
in the benzimidazole resistance of the small-ruminant parasite, Teladorsagia circumcincta. Parasitol. Res. 85,452–456.
Elard, L., Comes, A.M., Humbert, J.F., 1996. Sequences of -tubulin cDNA from benzimidazole-susceptible
and -resistant strains of Teladorsagia circumcincta, a nematode parasite of small ruminants. Mol. Biochem. Parasitol. 79, 249–253.
Elard, L., Sauvé, C., Humbert, J.F., 1998. Fitness of benzimidazole-resistant and -susceptible worms of
Teladorsagia circumcincta, a nematode parasite of small ruminants. Parasitology 117, 571–578.
Elard, L., Cabaret, J., Humbert, J.F., 1999. PCR diagnosis of benzimidazole-susceptibility or -resistance in natural
populations of the small ruminant parasite, Teladorsagia circumcincta. Vet. Parasitol. 80, 231–237.
Geary, T.G., Nulf, S.C., Favreau, M.A., Tang, L., Pritchard, R.K., Hatzenbuhler, N.T., Shea, M.H., Alexander, S.J.,
Klein, R.D., 1992. Three -tubulin cDNAs from the parasitic nematode Haemonchus contortus. Mol. Biochem. Parasitol. 50, 295–306.
Gettinby, G., Soutar, A., Armour, J., Evans, P., 1989. Anthelmintic resistance and the control of ovine ostertagiosis:
a drug action model for genetic selection. Int. J. Parasitol. 19, 369–376.
Grant, W.N., Mascord, L.J., 1996. -Tubulin polymorphism and benzimidazole resistance in Trichostrongyluscolubriformis. Int. J. Parasitol. 26, 71–77.
Hall, C.A., Ritchie, L., 1982. Effect of removing anthelmintic selection pressure on the benzimidazole resistance
status of Haemonchus contortus and Trichostrongylus colubriformis in sheep. Res. Vet. Sci. 33, 54–57.
Hennessy, D.R., Sangster, N., Steel, J.W., Collins, G.H., 1993. Comparative pharmacocinetic behaviour of
albendazole in sheep and goats. Int. J. Parasitol. 23, 321–325.
Hertzberg, H., Rossman, J., Kohler, L., Willi, U., 2000. Vorkommen von benzimidazol-resistenzen bei
magen-darmnematoden des schafes und der ziege in der Schweiz. Wien. Tierärztl. Mschr. 87, 3–9.
Hoste, H., Chartier, C., Etter, E., Goudeau, C., Soubirac, F., Lefrileux, Y., 2000. A questionnaire survey on the
practices adopted to control gastrointestinal nematode parasitism in dairy goat farms in France. Vet. Res. Commun. 24, 459–469.
Hubert, J., Kerboeuf, D., Nicolas, J.A., Dubost, G., Gayaud, G., 1991. Résistance des strongles gastro-intestinaux
aux benzimidazoles chez les petits ruminants en Limousin. Rec. Med. Vet. 167, 135–140.
Humbert, J.F., Elard, L., 1997. A simple method for rapidly detecting defined point mutations. Technical Tips
Online, http://tto.trends.com, T40076.
Kochapakdee, S., Pandey, V.S., Pralomkarn, W., Choldumrongkul, S., Ngampongsai, W., Lawpetchara, A., 1995.
Anthelmintic resistance in goats in southern Thailand. Vet. Rec. 137, 124–125.
Kwa, M.S., Kooyman, F.N.J., Boersema, J.H., Roos, M.H., 1993a. Effect of selection for benzimidazole resistance
in Haemonchus contortus in -tubulin isotype 1 and isotype 2 genes. Biochem. Biophys. Res. Commun. 191,413–419. J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
Kwa, M.S., Veenstra, J.G., Roos, M.H., 1993b. Molecular characterization of -tubulin genes present in
benzimidazole-resistant populations of Haemonchus contortus. Mol. Biochem. Parasitol. 60, 133–144.
Kwa, M.S., Veenstra, J.G., Roos, M.H., 1994. Benzimidazole resistance in Haemonchus contortus is correlated
with a conserved mutation at amino acid 200 in -tubulin isotype 1. Mol. Biochem. Parasitol. 63, 299–303.
Kwa, M.S., Veenstra, J.G., Dijk, M.D., Roos, M.H., 1995. -Tubulin genes from the parasitic nematode
Haemonchus contortus modulate drug resistance in Caenorhabditis elegans. J. Mol. Biol. 246, 500–510.
Leignel, V., 2000. Diversité génétique et résistance aux benzimidazoles chez Teladorsagia circumcincta
(Nematoda, Trichostrongylidae) parasite de petits ruminants. Ph.D. thesis, University of Montpellier 11(France), 157 pp.
Lubega, G.W., Klein, R.D., Geary, T.G., Prichard, R.K., 1994. Haemonchus contortus: the role of two -tubulin
gene subfamilies in the resistance to benzimidazole anthelmintics. Biochem. Pharmacol. 47, 1705–1715.
Maggs, A.F., Logan, J.M., Carter, P.E., Pennington, T.H., 1998. The detection of penicillin insensitivity in Neisseriameningitidis by polymerase chain reaction. J. Antimicrob. Chemother. 42, 303–307.
Maingi, N., Bjorn, H., Dangolla, A., 1998. The relationship between faecal egg count reduction and the lethal dose
50% in the egg hatch assay and larval development assay. Vet. Parasitol. 77, 133–145.
Marriner, S.E., Bogan, J.A., 1981. Anthelmintic efficacy. Vet. Rec. 109, 477–478. Martin, P.J., Anderson, N., Jarett, R.G., 1989. Detecting benzimidazole resistance with faecal egg count reduction
tests and in vitro assays. Aust. Vet. J. 66, 236–240.
O’Brien, T.F., 1997. The global epidemic nature of antimicrobial resistance and the need to monitor and manage
it locally. Clin. Infect. Dis. 24, s2–s8.
Presidente, P.J.A., 1985. Methods for detection of resistance to anthelmintics. In: Anderson, N., Waller, P.J. (Eds.),
Resistance in Nematodes to Anthelmintic Drugs. CSIRO, Division of Animal Health, Sydney, pp. 13–27.
Raymond, M., Callaghan, A., Fort, P., Pasteur, N., 1991. Worldwide migration of amplified insecticide resistance
genes in mosquitoes. Nature 350, 151–153.
Roos, M.H., Boersema, J.H., Borgsteede, F.H.M., Cornelissen, J., Taylor, M., Ruitenberg, E.J., 1990. Molecular
analysis of selection for benzimidazole resistance in the sheep parasite Haemonchus contortus. Mol. Biochem. Parasitol. 43, 77–88.
Silvestre, A., Chartier, C., Sauvé, C., Cabaret, J., 2000. Relationship between helminth species diversity, intensity
of infection and breeding management in dairy goats. Vet. Parasitol. 94, 91–105.
Silvestre, A., Humbert, J.F., 2000. A molecular tool for species identification and benzimidazole resistance
diagnosis in larval communities of small ruminant parasites. Exp. Parasitol. 95, 271–276.
Silvestre, A., 2000. Resistance aux benzimidazoles des communautés de nématodes parasites du tractus digestif
des petits ruminants: Mécanismes génétiques et factaurs environnementaux. Ph.D. Thesis, University of Touns(France), 79 pp.
Silvestre, A., Cabaret, J., Humbert, J.F., 2001. Effect of benzimidazole under-dosing on the resistant allele
frequency in Teladorsagia circumcincta (Nematoda). Parasitology 123, 103–111.
Smith, G., 1990. A mathematical model for the evolution of anthelmintic resistance in a direct life cycle nematode
parasite. Int. J. Parasitol. 20, 913–921.
Van Wyk, J.A., van Schalkwyk, P.C., 1990. A novel approach to the control of anthelmintic-resistant Haemonchuscontortus in sheep. Vet. Parasitol. 35, 61–69.
Van Wyk, J.A., Malan, F.S., Randles, J.L., 1997. How long before resistance makes it impossible to control some
field strains of Haemonchus contortus in South Africa with any of the modern anthelmintics? Vet. Parasitol. 70, 111–122.
Wood, I.B., Amaral, N.K., Bairden, K., Duncan, J.L., Kassai, T., Malone, J.B., Pankavich, J.A., Reinecke, R.K.,
Slocombe, O., Taylor, S.M., Vercruysse, J., 1995. World Association for the Advancement of VeterinaryParasitology (WAAVP) second edition of guidelines for evaluating the efficacy of anthelmintics in ruminants(bovine, ovine, caprine). Vet. Parasitol. 58, 181–213.
A SMOKE-FREE ZONE with NLP One year on since the smoking ban, T he ban on smoking in public places your health, plus public opinion granting Many smokers have mixed feelings about it honorary pariah status, combined with quitting: they want to lose the disadvantages the current legislation making it so darned of smoking but don’t want to lose the benefits. polls, ‘three quarters of
Approximate seed count (raw): 7,400 to 7,600 Soil temperature: 71 to 73°F (21 to 22°C) Light: Up to 2,500 f.c. (26,900 Lux) Moisture: Reduce soil moisture slightly (level 3 Key flowering facts: to 4) to allow the roots to penetrate into the • Vernalization: not required but beneficial as Fertilizer: Apply fertilizer at rate 1 (less than 100 flowering will occur 2 to

 Veterinary Parasitology 101 (2001) 405–414
Molecular approaches to studying benzimidazole
J.F. Humbert a,∗, J. Cabaret b, L. Elard c, V. Leignel d, A. Silvestre b
a INRA, Station d’Hydrobiologie Lacustre, BP 511, 74203 Thonon Cedex, France
b INRA, Station de Pathologie Aviaire et de Parasitologie, 37380 Nouzilly, France
c Laboratoire de Parasitologie Fondamentale et Fonctionnelle,
Université de Paris VI, 75252 Paris Cedex 05, France
d Laborato´ıre de Biolog´ıe et de Génétique Evolutive, Faculté des Sciences et Techniques,
Abstract
Veterinary Parasitology 101 (2001) 405–414
Molecular approaches to studying benzimidazole
J.F. Humbert a,∗, J. Cabaret b, L. Elard c, V. Leignel d, A. Silvestre b
a INRA, Station d’Hydrobiologie Lacustre, BP 511, 74203 Thonon Cedex, France
b INRA, Station de Pathologie Aviaire et de Parasitologie, 37380 Nouzilly, France
c Laboratoire de Parasitologie Fondamentale et Fonctionnelle,
Université de Paris VI, 75252 Paris Cedex 05, France
d Laborato´ıre de Biolog´ıe et de Génétique Evolutive, Faculté des Sciences et Techniques,
Abstract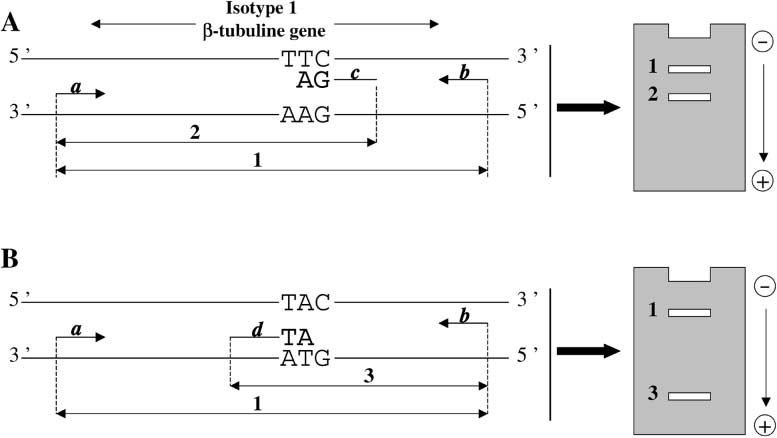
 J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
Fig. 1. Principle of allele-specific PCR for the genotyping of worms in respect of residue 200 of their isotype 1-tubulin gene: (A) detection of BZ-susceptible allele; (B) detection of BZ-resistant allele. This allele-specificPCR contains four primers: a and b are BZ-non-specific primers; c is a BZ-susceptible primer; d is a BZ-resistantprimer. Three fragments can thus be amplified: a BZ-non-specific fragment (1); a BZ-susceptible fragment (2); and aBZ-resistant fragment (3). In homozygous TTC/TTC worms only two fragments (1 and 2) are detected in agaroseelectrophoresis. In homozygous TAC/TAC worms, only two fragments (2 and 3) are detected in agarose elec-trophoresis. In heterozygous TTC/TAC worms, the three fragments (1–3) are detected in agarose electrophoresis.
J.F. Humbert et al. / Veterinary Parasitology 101 (2001) 405–414
Fig. 1. Principle of allele-specific PCR for the genotyping of worms in respect of residue 200 of their isotype 1-tubulin gene: (A) detection of BZ-susceptible allele; (B) detection of BZ-resistant allele. This allele-specificPCR contains four primers: a and b are BZ-non-specific primers; c is a BZ-susceptible primer; d is a BZ-resistantprimer. Three fragments can thus be amplified: a BZ-non-specific fragment (1); a BZ-susceptible fragment (2); and aBZ-resistant fragment (3). In homozygous TTC/TTC worms only two fragments (1 and 2) are detected in agaroseelectrophoresis. In homozygous TAC/TAC worms, only two fragments (2 and 3) are detected in agarose elec-trophoresis. In heterozygous TTC/TAC worms, the three fragments (1–3) are detected in agarose electrophoresis.