Scientia Horticulturae 105 (2005) 263–268
Anthony Koutoulis ,Ashis T. Roy Aina Price
a Cell Biology Group, School of Plant Science, University of Tasmania,
Private Bag 55, Hobart, Tasmania 7001, Australia
b Australian Hop Marketers, GPO Box 104A, Hobart, Tasmania 7001, Australia
Received 9 December 2003; received in revised form 20 July 2004; accepted 20 January 2005
The genome doubling agent colchicine has been used effectively to obtain tetraploid plants when
starting with diploid material. Since the plant meristem consists of many cells, as well as obtainingtetraploids, it is possible to obtain mixoploids (chimeras consisting of diploid and tetraploid tissue)after colchicine treatment. Flow cytometry was used to determine the DNA ploidy level of differenttissues of individual hop plants following colchicine treatment of diploid hops. Plants that are shownto be mixoploid after analysis of leaf tissue can display higher levels of tetraploid nuclei in root tissue,such that they could be mistakenly classified as tetraploid. The hop variety Galena-4n, which waspreviously considered to be a tetraploid, is shown to be a mixoploid based on leaf material. However,Galena-4n consistently produces triploid progeny when crossed with a diploid hop variety, indicatingthat Galena-4n produces tetraploid reproductive tissues and is effectively a tetraploid from a breedingperspective. The results from this study indicate that care should be taken when determining the DNAploidy level of an individual plant after exposure to genome-doubling agents based on single tissuesand that leaf tissue is a choice material for flow cytometric analysis. # 2005 Elsevier B.V. All rights reserved.
Keywords: Chimera; Colchicine; Flow cytometry; Hop; Mixoploid; Tetraploid
* Corresponding author. Tel.: +61 3 62262737; fax: +61 3 62262698.
E-mail address: anthony.koutoulis@utas.edu.au (A. Koutoulis).
0304-4238/$ – see front matter # 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.scienta.2005.01.010
A. Koutoulis et al. / Scientia Horticulturae 105 (2005) 263–268
Hop (Humulus lupulus L.) is a dioecious perennial climbing plant (During
winter, the plant goes into a dormant phase; a rootstock remains, which consists of stem andperennial root tissue During spring, buds on the rootstock grow to formthe climbing stems or ‘‘hop bines’’, which give rise to leaf and reproductive tissues. Theinflorescences of the female hop are of great importance to the brewing industry as theycontain the resins and essential oils that impart bitterness and aroma to beer ). Tetraploid induction has been utilized in anumber of breeding programs for the production of new varieties. In Australia, tetraploidfemale hops play an important role, as they can be crossed with diploid male hops toproduce triploid progeny. In Australian conditions, triploid hops are superior as they aremore vigorous, higher yielding and relatively seedless (unpublished data).
In previous work, we described an efficient in vitro colchicine-mediated technique for
the induction of tetraploid hops; a technique that can also generate mixoploid hops DNA ploidy determination was based on flow cytometric analysis of nucleiisolated from leaf tissue. This paper reports on the DNA ploidy level of nuclei isolated fromdifferent tissues from individual hop plants using flow cytometry and demonstrates thatmixoploids may be useful in a breeding program.
The diploid hop variety H138, in vitro colchicine-treated plants derived from it from
previous work (and Galena-4n were investigated. Galena-4n is a female,putative tetraploid, imported to Australia from the USA, generated from the diploid Galenahop variety following ex vitro colchicine treatment (G. Probasco, pers. comm.). Growthconditions (in vitro and glasshouse) are described in Tissue types ofplants (1–3 cm high) tested included: (i) leaf (the youngest, expanded leaf); (ii) shoot apex(the unexpanded tip); (iii) stem (the internode above the youngest expanded leaf); and (iv)root (the first 10 mm of root tips).
For flow cytometry analysis nuclei were collected by chopping 1 g of leaf material in
Galbraith’s buffer (on ice, stained with 1 mg lÀ1 propidium iodideand analysed using a Coulter Elite ESP flow cytometer equipped with a 100 W high-pressure mercury arc lamp (excitation using a 488 nm air cooled argon laser andflourescence collected using a 600 nm dichroic LP and 575 nm BP) as described in Bean (Vicia faba) nuclei were used as an internal standard.
Flow cytometric analysis produces histograms of DNA intensity, which is related to
DNA ploidy level (Samples were classed into ploidy types according to the peaksobtained. In addition to diploid (top) and tetraploid bottom), there weremixoploid (a chimera of diploid and tetraploid nuclei) classes , middle). Diploids
A. Koutoulis et al. / Scientia Horticulturae 105 (2005) 263–268
Fig. 1. Representative flow cytometer histograms (bean internal control peak not included). (Top) Diploid profileshowing diploid (2n) and tetraploid (4n) peaks. (Middle) Mixoploid profile showing diploid (2n), tetraploid (4n)and octaploid (8n) peaks. (Bottom) Tetraploid profile showing tetraploid (4n) and octaploid (8n) peaks.
A. Koutoulis et al. / Scientia Horticulturae 105 (2005) 263–268
Table 1DNA ploidy type by tissue of Galena-4n and examples of individual H138-derived in vitro hop plants
* Classification of individual plants was based on results obtained from analysis of leaf tissue (i.e., a diploid
plant has a diploid profile of leaf material but may have a different ploidy profile in other tissues) (Mixo-G1 = grade-1 mixoploid; Mixo-G2 = grade-2 mixoploid; Mixo-G3 = grade-3 mixoploid).
** The root sample of Galena-4n had some debris associated with the diploid peak. Consequently, this value may
slightly overestimate the percentage of diploid nuclei; however, the high percentage of tetraploid and octoploidnuclei indicates a significant amount of tetraploid cells in the root sample of Galena-4n.
possessed a small percentage of nuclei with a tetraploid complement of DNA (top),which represents nuclei at the late S or G2 phase of the cell cycle. Similarly, mixoploids(middle) and tetraploids (bottom) possessed octaploid nuclei.
The percentage of nuclei within each ploidy peak (diploid, tetraploid or octaploid) can
be calculated according to the number of counts within each peak. These results were usedto classify individual plants and/or tissues (see ). Mixoploids were graded accordingto the relative amounts of diploid and tetraploid nuclei; grade-1 mixoploids had morediploid than tetraploid nuclei, grade-2 mixoploids had similar amounts of diploid andtetraploid nuclei and grade-3 mixoploids had less diploid than tetraploid nuclei.
From previous work (), H138-derived hop plants with a range of DNA
ploidy types were available (diploid, grade-1 mixoploid, grade-2 mixoploid, grade-3mixoploid and tetraploid). In this study, the initial DNA ploidy level of a plant was based onthe result obtained from flow cytometry testing of leaf material. We determined the DNAploidy level of different tissue types of individual plants with different ploidy types(Leaf and shoot apex tissue produced similar ploidy profiles within an individualplant, while results from stem and root tissue conferred higher ploidy levels than leaf andshoot apex tissue within an individual plant (This was more pronounced in roottissue and can be attributed to the higher number of cells undergoing cell division in theroot. Importantly, although a grade-3 mixoploid could be separated from a tetraploid usingleaf tissue, it was not possible to differentiate between a grade-3 mixoploid and a tetraploidusing root tissue After testing leaf material using flow cytometry, Galena-4n wasclassified as a mixoploid (). When root tissue of Galena-4n was analysed using flowcytometry, the sample exhibited a tetraploid profile
A. Koutoulis et al. / Scientia Horticulturae 105 (2005) 263–268
The results of this study show that a grade-3 mixoploid could be separated from a
tetraploid using leaf tissue; however, it was not possible to differentiate between a grade-3mixoploid and a tetraploid using root tissue. Furthermore, this study shows that the putativetetraploid Galena-4n possesses a mixoploid flow cytometry profile when using leaf materialand a tetraploid flow cytometry profile when using root material. This plant was identified as atetraploid by chromosome counts of root tissue (G. Probasco, pers. comm.). This casts doubton the reliability of using root tissue for DNA ploidy testing via flow cytometry.
Interestingly, triploid progeny have been produced from Galena-4n when crossed with a
male diploid in the USA and Australia (unpublished data). As Galena-4n has been shown tobe a mixoploid in this study, the breeding data suggests that some or all of the femalereproductive tissues of Galena-4n are tetraploid. In the 2002/2003 Australian breedingseason, when Galena-4n was crossed to three different diploid males under controlled crossconditions (i.e., covered with sleeves), 8 of the 9 (3 from each cross) progeny tested by flowcytometry were triploid while the remaining one was tetraploid. At this stage it is unclearhow a tetraploid could arise from Galena-4n when crossed with a male diploid undercontrolled conditions, however, possibilities include parthenogenesis or an unreduceddiploid male gamete. In the 2000/2001 Australian breeding season, 11 of 12 progeny testedby flow cytometry from open pollinated Galena-4n plants were triploid while theremaining one was tetraploid. Again parthenogenesis could account for the tetraploid,however, as it was open-pollinated, alternative explanations include an unreduced diploidmale gamete from a diploid male or a normally reduced diploid male gamete from atetraploid male present in the hop garden.
As Galena-4n can produce triploid progeny when crossed with a diploid, our data
indicates that mixoploid hops can play a role in a triploid breeding program. It is difficult totest the DNA ploidy level of female reproductive tissue in a mixoploid via flow cytometry,as only a very small part of the flower, the egg within the embryo sac, contributes to theploidy level of the progeny. An alternative method to evaluate the ploidy of the egg is todirectly assess the seedlings produced. This will form part of our future work and it willallow us to determine whether separating mixoploids into different grades has any practicalvalue with respect to female reproductive parts.
Caution should be used when investigating colchicine-derived mixoploids, due to the
biology of the hop plant. After a dormant phase in winter, buds on the rootstock grow toform the hop bines during spring, which give rise to leaf and reproductive tissues (Presently, we have no knowledge of the meristematic regions of these mixoploidhops and do not know whether they represent periclinal chimeras of the type used in theelegant work by which demonstrated the three germ layers (LI, LII andLIII) in the shoot apex of Datura. Subsequent work by showed that the LIIlayer in Datura gives rise to the nucellus (and embryo sac) within the ovule. If thisphenomenon holds true in hops, then periclinal hop chimeras with a tetraploid LII layercould be used in the triploid breeding program. Interestingly, the periclinal grape (Vitisvinifera) chimera (Pinot Meunier) has been developed into an agronomically importantcultivar ). At this stage our results indicate thatGalena-4n is a stable mixoploid that is effectively a tetraploid for breeding purposes.
A. Koutoulis et al. / Scientia Horticulturae 105 (2005) 263–268
It is possible that the other generated mixoploid hops are not stable and that the ratio of
diploid and tetraploid cells is different in different bines originating from the samerootstock. If this were the case and if a relationship was found between mixoploid gradeand ability to form tetraploid female reproductive parts, mixoploid hop bines would need tobe graded each year; far from an ideal situation. However, if required, it is possible togenerate a new pure tetraploid plant by using an in vitro callus-based technique, describedin , using a mixoploid as starting material.
This study demonstrates that the type of tissue used for flow cytometry analysis can
impact on the results obtained. Where the DNA ploidy level of a plant needs to beaccurately determined the leaf would be the best source of testing material.
We thank Mr. Mark Cozens for assistance with flow cytometry. Funding from Australian
Hop Marketers and Horticulture Australia supported this work.
Boss, P.K., Thomas, M.R., 2002. Association of dwarfism and floral induction with a grape ‘green revolution’
Burgess, A.H., 1964. Hops: Botany, Cultivation and Utilization. Interscience Publishers, New York. Franks, T., Botta, R., Thomas, M.R., 2002. Chimerism in grapevines: implications for cultivar identity, ancestry
and genetic improvement. Theor. Appl. Genetics 104, 192–199.
Galbraith, D.W., Harkins, K.R., Maddox, J.M., Ayres, N.M., Sharma, D.P., Firoozabady, E., 1983. Rapid flow
cytometric analysis of the cell cycle in intact plant tissues. Science 220, 149–151.
Haunold, A., 1980. Hop. In: Fehr, W.R., Hadley, H.H. (Eds.), Hybridization of Crop Plants. American Society of
Agronomy, Madison, WI, pp. 393–406.
Neve, R.A., 1991. Hops. Chapman and Hall, London. Roy, A.T., Leggett, G., Koutoulis, A., 2001a. In vitro tetraploid induction and generation of tetraploids from
mixoploid in hop (Humulus lupulus L.). Plant Cell Reports 20, 489–495.
Roy, A.T., Leggett, G., Koutoulis, A., 2001b. Development of a shoot multiplication system for hop (Humulus
lupulus L.). In Vitro Cell. Dev. Biol.: Plant 37, 79–83.
Satina, S., 1945. Periclinal chimeras in Datura in relation to the development and structure of the ovule. Am. J.
Satina, S., Blakeslee, A.F., Avery, A.G., 1940. Demonstration of the three germ layers in the shoot apex of Datura
by means of induced polyploidy in periclinal chimeras. Am. J. Botany 27, 895–905.
Verzele, M., De Keukeleire, D., 1991. Chemistry and analysis of hop and beer bitter acids. Developments in Food
Science, vol. 27. Elsevier, Amsterdam.
A. Biological Chemistry(though water is the universal medium for life on earth, living organisms are made of chemicals based mostly on the element a. contain Carbon. total of 6e-, 2 in 1st shel , 4 in 2nd that holds 8 i. Carbon has 4 valence electrons that can join with an electron from another atom to form a strong covalent bond-usual y with C, H, O, or N ex/ CH4 (methane) ii. Carbon can
Press Release Press contact: Corporate C ommunications Office Tel: +81-3-3457-2105 1-1-1 Shibaura, Minato-ku, Tokyo 105-8001, Japan Fax: +81-3-5444-9202 URL: http://www.toshiba.co.jp/about/press/index.htm Toshiba Signs Agreement to Develop Next Generation Thermal Power System with NET Power, Shaw and Exelon - To develop an innovative thermal power system


 Scientia Horticulturae 105 (2005) 263–268
Anthony Koutoulis ,Ashis T. Roy Aina Price
a Cell Biology Group, School of Plant Science, University of Tasmania,
Private Bag 55, Hobart, Tasmania 7001, Australia
b Australian Hop Marketers, GPO Box 104A, Hobart, Tasmania 7001, Australia
Received 9 December 2003; received in revised form 20 July 2004; accepted 20 January 2005
The genome doubling agent colchicine has been used effectively to obtain tetraploid plants when
starting with diploid material. Since the plant meristem consists of many cells, as well as obtainingtetraploids, it is possible to obtain mixoploids (chimeras consisting of diploid and tetraploid tissue)after colchicine treatment. Flow cytometry was used to determine the DNA ploidy level of differenttissues of individual hop plants following colchicine treatment of diploid hops. Plants that are shownto be mixoploid after analysis of leaf tissue can display higher levels of tetraploid nuclei in root tissue,such that they could be mistakenly classified as tetraploid. The hop variety Galena-4n, which waspreviously considered to be a tetraploid, is shown to be a mixoploid based on leaf material. However,Galena-4n consistently produces triploid progeny when crossed with a diploid hop variety, indicatingthat Galena-4n produces tetraploid reproductive tissues and is effectively a tetraploid from a breedingperspective. The results from this study indicate that care should be taken when determining the DNAploidy level of an individual plant after exposure to genome-doubling agents based on single tissuesand that leaf tissue is a choice material for flow cytometric analysis.
Scientia Horticulturae 105 (2005) 263–268
Anthony Koutoulis ,Ashis T. Roy Aina Price
a Cell Biology Group, School of Plant Science, University of Tasmania,
Private Bag 55, Hobart, Tasmania 7001, Australia
b Australian Hop Marketers, GPO Box 104A, Hobart, Tasmania 7001, Australia
Received 9 December 2003; received in revised form 20 July 2004; accepted 20 January 2005
The genome doubling agent colchicine has been used effectively to obtain tetraploid plants when
starting with diploid material. Since the plant meristem consists of many cells, as well as obtainingtetraploids, it is possible to obtain mixoploids (chimeras consisting of diploid and tetraploid tissue)after colchicine treatment. Flow cytometry was used to determine the DNA ploidy level of differenttissues of individual hop plants following colchicine treatment of diploid hops. Plants that are shownto be mixoploid after analysis of leaf tissue can display higher levels of tetraploid nuclei in root tissue,such that they could be mistakenly classified as tetraploid. The hop variety Galena-4n, which waspreviously considered to be a tetraploid, is shown to be a mixoploid based on leaf material. However,Galena-4n consistently produces triploid progeny when crossed with a diploid hop variety, indicatingthat Galena-4n produces tetraploid reproductive tissues and is effectively a tetraploid from a breedingperspective. The results from this study indicate that care should be taken when determining the DNAploidy level of an individual plant after exposure to genome-doubling agents based on single tissuesand that leaf tissue is a choice material for flow cytometric analysis.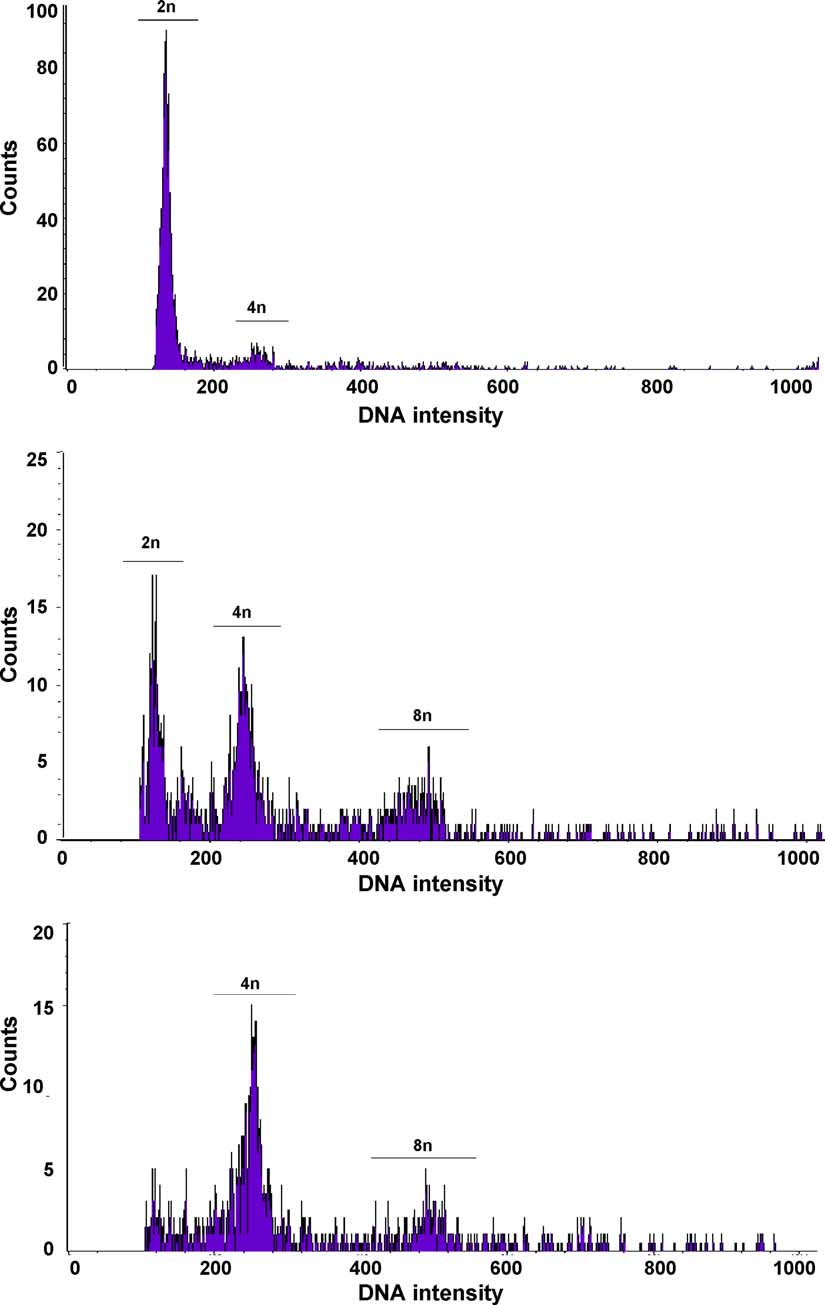 A. Koutoulis et al. / Scientia Horticulturae 105 (2005) 263–268
Fig. 1. Representative flow cytometer histograms (bean internal control peak not included). (Top) Diploid profileshowing diploid (2n) and tetraploid (4n) peaks. (Middle) Mixoploid profile showing diploid (2n), tetraploid (4n)and octaploid (8n) peaks. (Bottom) Tetraploid profile showing tetraploid (4n) and octaploid (8n) peaks.
A. Koutoulis et al. / Scientia Horticulturae 105 (2005) 263–268
Fig. 1. Representative flow cytometer histograms (bean internal control peak not included). (Top) Diploid profileshowing diploid (2n) and tetraploid (4n) peaks. (Middle) Mixoploid profile showing diploid (2n), tetraploid (4n)and octaploid (8n) peaks. (Bottom) Tetraploid profile showing tetraploid (4n) and octaploid (8n) peaks.